Царство растений
Биология — Тейлор Д., Грин Н., Стаут У. 1 том — 2013
Хотя жизнь на нашей планете, вероятно, зародилась примерно 3,5 млрд. лет назад, первые организмы заселили сушу не ранее, чем 420 млн. лет назад. Это были самые первые растения. Растения — автотрофные эукариоты, которые адаптировались к жизни в воздушной среде. Единственные другие автотрофы среди эукариот — это водоросли, специализировавшиеся к жизни в воде. Напоминаем: автотрофами называют организмы, использующие в качестве источника для синтеза органических веществ неорганический углерод, а именно диоксид углерода. Однако помимо источника углерода для такого синтеза требуется еще и энергия, и растения, будучи фотоавтотрофами, используют в качестве источника энергии свет. Этот способ питания более принято называть фотосинтезом.
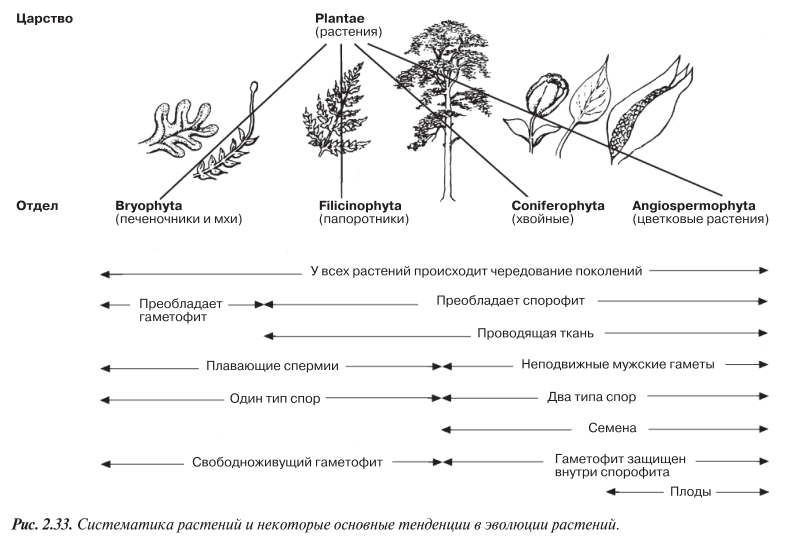
История эволюции растений — это постепенно совершенствующаяся адаптация к жизни на суше. Именно эта история и составит одну из основных тем в нашем изучении растений. Систематика растений, показана на рис. 2.33. Там же приводится краткий перечень некоторых основных тенденций в эволюции растений, связанных с адаптацией к жизни на суше, которые также будут рассматриваться в этом разделе.
2.7.1. Отдел Bryophyta (печеночники и мхи)
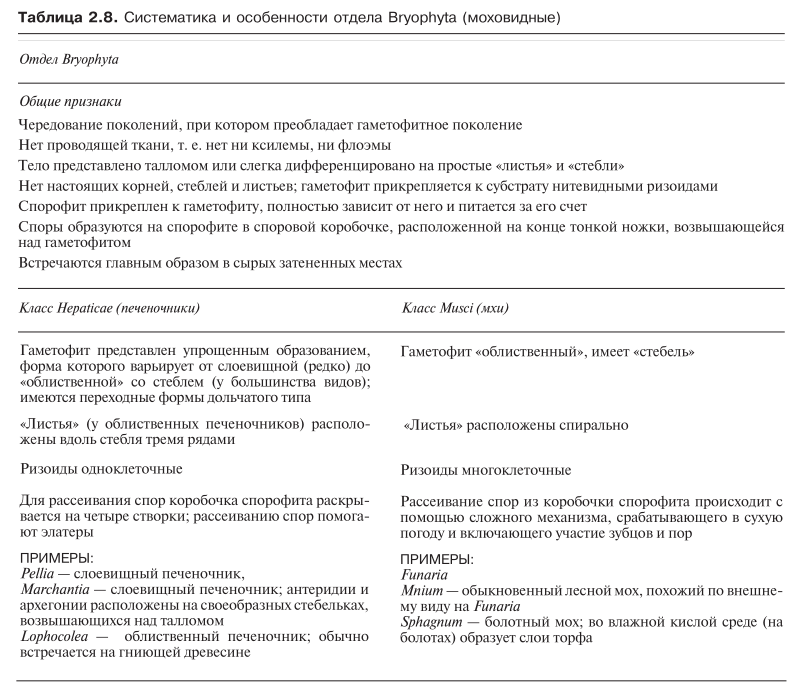
Печеночники и мхи — наиболее примитивные из всех наземных растений. Полагают, что они произошли от зеленых водорослей. В отдел Bryоphyta (мохообразные) входят два главных класса — Hepaticae (печеночники) и Musci (мхи). Обе эти группы плохо приспособлены к жизни на суше, и поэтому они привязаны к сырым затененным местам. Систематика и основные признаки мохообразных приведены в табл. 2.8.
Bryophyta — небольшие растения с несложным строением. Опорная и проводящая ткани у них развиты слабо или же вовсе отсутствуют. Нет у них дифференцированной ксилемы и флоэмы, так же как нет и настоящих корней. В почве они удерживаются тончайшими нитями, называемыми ризоидами. Вода и минеральные соли поглощаются всей поверхностью тела, в том числе и ризоидами. А это означает, что в отличие от настоящих корней ризоиды служат лишь для закрепления растения в грунте. (В на стоящих корнях, как и в настоящих стеблях или листьях имеются проводящие ткани.) Мохообразные не покрыты сверху кутикулой, либо кутикула столь тонка, что она не препятствует потере (или поступлению) воды. Тем не менее многие мохообразные приспособились выдерживать засушливые периоды, используя для этой цели какие-то не совсем понятные механизмы. Например, было установлено, что такой хорошо известный ксерофитный мох, как Grimmia pulvinata, больше года остается живым при 20 °С в абсолютно высушенном состоянии.
Сразу же после того, как растение попадает во влажную среду, у него восстанавливаются все функции.
Чередование поколений
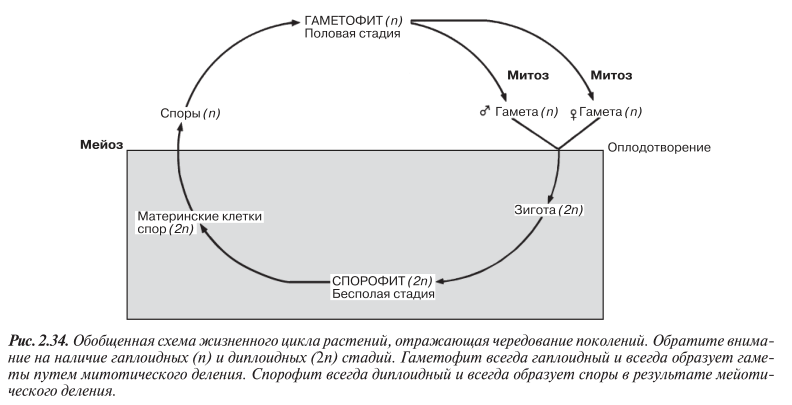
Как у всех наземных растений и некоторых наиболее высоко организованных водорослей, таких как ламинария, у мохообразных наблюдается чередование поколений. В ходе жизненного цикла происходит смена двух типов организмов: поколение гаплоидного гаметофита и поколение диплоидного спорофита поочередно сменяют друг друга, что схематически показано на рис. 2.34. Гаплидное поколение называется гаметофитом (от греч. gametе — жена, gametеs — муж; phyton — растение), так как оно способно к половому размножению и образует гаметы. Поскольку образование гамет происходит в результате митоза, они тоже гаплоидны. Сливаясь, гаметы образуют диплоидную зиготу, из которой вырастает следующее поколение — поколение диплоидных спорофитов. Они называются спорофитами, потому что способны к бесполому размножению с образованием спор. Споры об разуются в результате мейоза, т. е. здесь происходит возврат к гаплоидному состоянию. Гаплоидные споры дают начало гаметофитному поколению. Одно из этих двух поколений всегда преобладает над другим, и на его долю приходится большая часть жизненного цикла; это поколение называют доминантным. У моховидных до минирует поколение гаметофитов, у всех остальных наземных растений — поколение спорофитов. Доминирующее поколение принято помещать в верхнюю половину схемы, изображающей жизненный цикл.
На рис. 2.34 в обобщенном виде представлен жизненный цикл всех наземных растений, в том числе и наиболее высокоорганизованных цветковых растений. Никогда не забывайте, что гаметы у растений образуются не в результате мейоза, как у животных, а в результате митоза; мейотическое деление происходит при образовании спор.
Класс Hepaticae — печеночники
Характерные признаки класса Hepaticae представлены в табл. 2.8. По своему строению печеночники намного проще, чем мхи, и в целом они более привязаны к сырым и затененным местам.
Их можно найти на берегу рек и ручьев, на влажных камнях и среди болотной растительности. У большинства печеночников видны правильные доли или вполне выраженные «стебли» с небольшими простыми «листочками». Проще всего устроены талломные печеночники, тело которых представлено плоским слоевищем и не разделе но на стебли и листья.
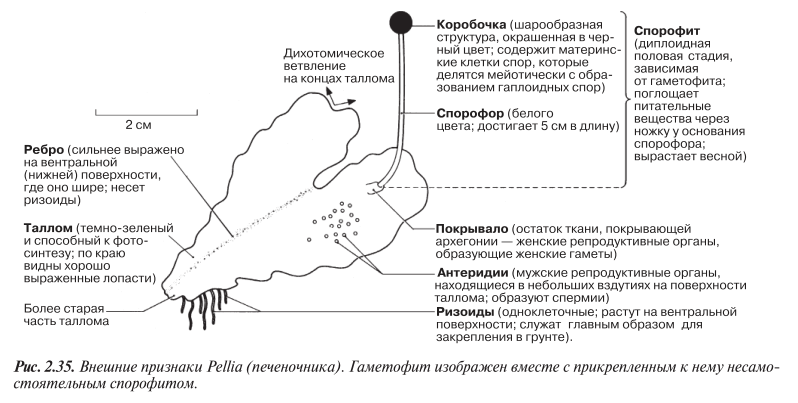
Примером может служить Pellia, печеночник, широко распространенный по всей Великобритании. Это растение окрашено в тускло-зеленый цвет, ширина плоских «веточек» составляет около 1 см. Внешние признаки Pellia приведены на рис. 2.35.
Класс Musci — листостебельные мхи
Основные признаки листостебельных мхов перечислены в табл. 2.8. Листостебельные мхи гораздо более дифференцированы, чем печеночники, но, как и печеночники, это небольшие растения, встречающиеся главным образом в сырых местах. Они часто образуют плотные по душки.
Funaria — обычный представитель лиственных мхов на полях, вырубках и перекопанных землях, где она поселяется одной из первых. Funaria особенно любит селиться на кострищах и пожарищах. Это один из самых обычных сорняков в теплицах и садах. Внешний вид Funaria по казан на рис. 2.36.
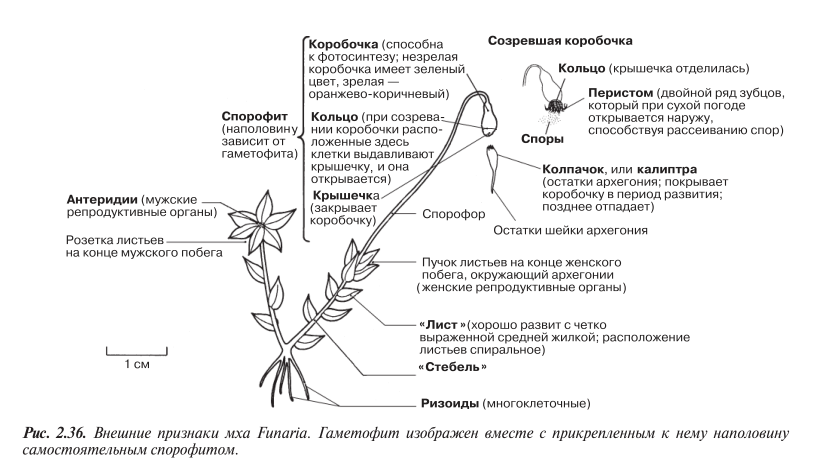
Как и у печеночников, для оплодотворения Funaria необходима вода. Когда поверхность таллома увлажняется, созревшие антеридии поглощают воду и лопаются, высвобождая на поверхность мужские гаметы (спермии). Каждый спермий снабжен двумя жгутиками. Спермии подплывают к архегониям, в каждом из которых находится женская гамета, или яйцеклетка. Оплодотворение, т. е. слияние ядра спермия с яд ром яйцеклетки, происходит в архегонии. В результате слияния образуется диплоидная зигота, которая, вырастая из архегония, дает начало новому спорофиту.
2.7.2. Отдел Filicinophyta (папоротниковидные)
Основные признаки Filicinophyta перечислены в табл. 2.9. Папоротники обычно встречаются только в тенистых влажных местах. Немногие папоротники могут расти на открытом месте, хотя самый обычный орляк (Pteridium) составляет исключение. Папоротники широко распространены в тропических дождевых лесах, где температура, освещенность и влажность наиболее благоприятны для них.

Папоротники — растения сосудистые; иными словами у них имеется проводящая ткань, которая состоит из ксилемы и флоэмы, выполняющих функцию транслокации (пере носа) воды и питательных веществ в теле растения. По сосудам ксилемы осуществляется перенос в основном воды и минеральных солей, тогда как флоэма переносит главным образом растворы органических веществ, таких как сахара. Проводящие ткани являются серьезным эволюционным преимуществом по сравнению с простыми проводящими клетками некоторых моховидных и водорослей. Эти ткани обнаружены только в спорофитном поколении и именно поэтому спорофиты становятся доминирующими у всех сосудистых растений.
Проводящие ткани выполняют две важные функции. Во-первых они образуют транспортную систему, переносящую питательные вещества и воду по многоклеточному телу, обеспечивая возможность развития крупного сложного тела. Во-вторых, они выполняют и опорную функцию, поскольку ксилема, будучи проводящей тканью, содержит лигнифицированные клетки — очень прочные и твердые. У сосудистых растений развивается и другая лигнифицированная ткань — склеренхима. Она усиливает механическую роль ксилемы.
Спорофитное поколение имеет настоящие корни, стебли и листья. Корни проникают в почву, что облегчает поступление в растение воды и растворенных веществ. По ксилеме они поступают в другие части растения.
Как только тело растения, получив поддержку, смогло возвыситься над поверхностью земли, сразу же должна была возникнуть конкуренция за свет, и в результате появилась тенденция к развитию все более высоких форм.
Папоротники и древовидные папоротники доминировали на суше в течение примерно 70 млн. лет, начиная с девонского периода до пермского. Затем они были в значительной степени вытеснены сначала хвойными, а позднее и цветковыми растениями.
Несмотря на существенные прогрессивные адаптации спорофитного поколения к воздушной среде, гаметофит папоротниковидных, называемый заростком, все еще сталкивался с большими проблемами. Он еще меньше по раз мерам и менее стоек к обезвоживанию, чем гаметофит у моховидных. На заростках образуются спермии, которые, как и у моховидных, могут достичь женских гамет, только подплыв к ним.
Щитовник мужской (Dryopteris filix-mas)
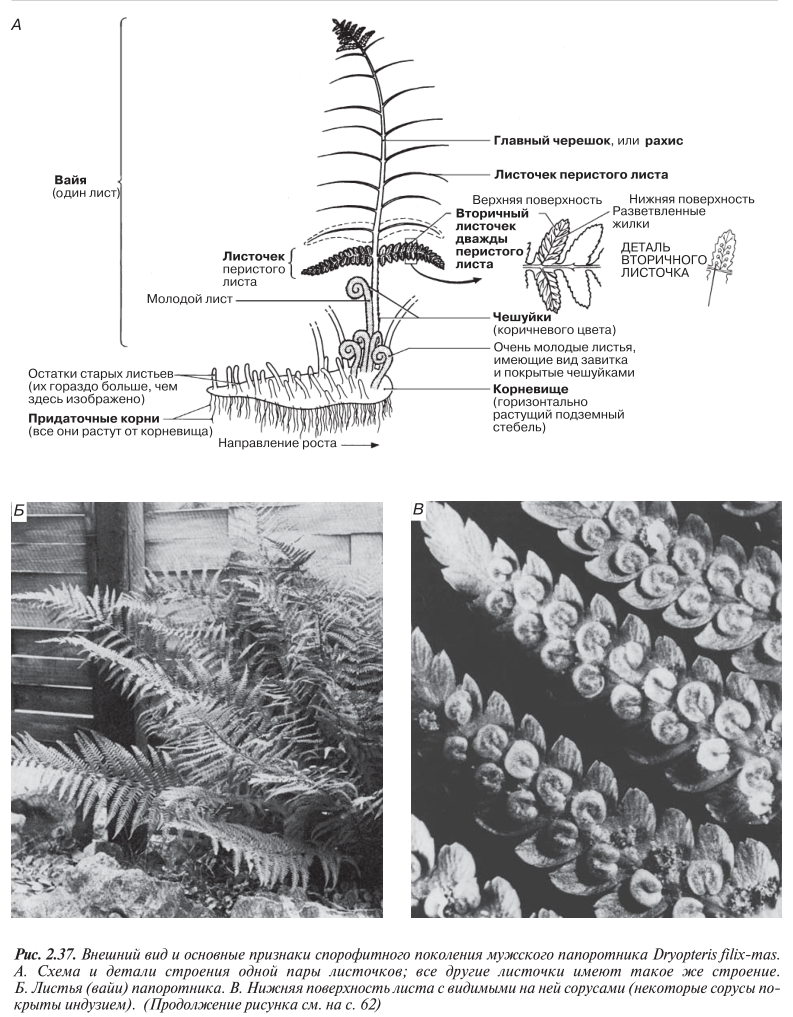
Это, пожалуй, самый распространенный в Великобритании папоротник; он встречается по всей стране в сырых лесах, лесопарках и других тенистых местах. Вайи (листья) спорофита, достигающие в высоту до 1 м и более, рас тут от толстого горизонтального стебля, или корневища. На корневище находятся придаточные корни. От основного корневища могут отламываться отдельные ветви и давать начало новым растениям. Это — одна из форм вегетативного размножения. У основания корневище покрыто сухими бурыми чешуйками, защищающими молодые листья от заморозков и от засухи. Молодые листья плотно закручены в характерные для папоротников завитки. Выше на черешке листа размеры чешуек постепенно уменьшаются, и расстояния между ними увеличиваются. Черешок вайи называется главным черешком, а листочки, отходящие от него в обе стороны, — листочками перистого листа. Небольшие округлые выступы на листочках называются вторичным листочками. С внешними признаками спорофита Dryopteris filix-mas можно познакомиться на рис. 2.37; спорофит показан на рис. 2.38.
Споры образуются в конце лета в специальных структурах, называемых спорангиями.
Спорангии находятся на нижней стороне вторичных листочков в особых скоплениях, получивших название сорусов (рис. 2.37, В, Г и Д). Каждый сорус покрыт защитным покрывалом — индузием. Внутри каждого спорангия происходит мейотическое деление диплоидных материнских клеток спор с образованием в результате гаплоидных спор. Созрев, индузий сморщивается и отпадает, а открывшиеся стенки спорангия начинают подсыхать. В конце концов стенка разрывается и споры «выстреливают» из спорангия как из катапульты (рис. 2.37, Д).
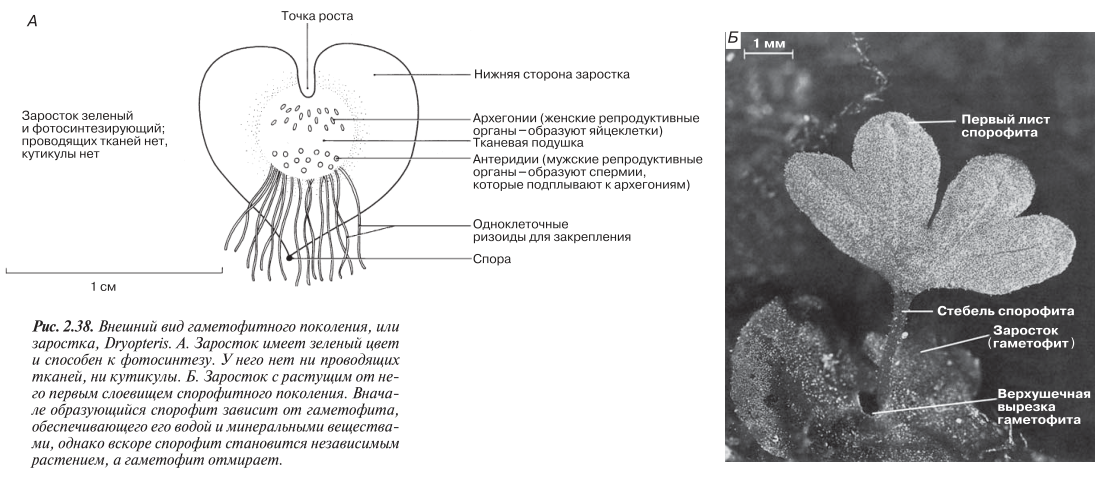
Споры прорастают, давая начало гаметофитному поколению. Гаметофит представляет собой тонкую сердцевидную пластинку клеток диаметром около 1 см (рис. 2.38). Эта пластинка имеет зеленый цвет, способна к фотосинтезу и прикрепляется к почве одноклеточными ризоидами. Поскольку у такого нежного заростка нет кутикулы, он быстро высыхает и, следовательно, может жить только во влажной среде.
На нижней стороне гаметофита (заростка) образуются простые архегонии и антеридии.
Эти репродуктивные органы защищают находящиеся в них гаметы. Гаметы возникают в результате митоза из материнских половых клеток; при этом, как и у моховидных, в антеридиях образуются спермии, а в каждом архегонии — по одной яйцеклетке. Спермии снабжены жгутиками. Во влажных условиях созревшие спермии высвобождаются из антеридиев и по водной пленке подплывают к архегониям. В результате оплодотворения образуется диплоидная зигота. Обратите внимание, что оплодотворение у папоротниковидных, также как и у моховидных, все еще зависит от наличия воды.
Зиготы дают начало спорофитному поколению. Молодой зародыш поглощает питательные вещества из гаметофита до тех пор, пока эту функцию не возьмут на себя собственные листья и корни (рис. 2.38, Б). Гаметофит вскоре увядает и отмирает.
2.7.3. Семенные растения
Самая процветающая группа растений образует семена. Эти растения, по-видимому, произошли от вымерших семенных представителей папоротников и их близких родичей. Систематика и основные признаки семенных растений суммированы в табл. 2.10.


В табл. 2.10 рассматриваются две основные группы семенных растений — голосеменные и покрытосеменные. Последние чаще называют цветковыми растениями. У голосеменных семязачатки (и впоследствии семена) располагаются на поверхности особых чешуйчатых листьев, называемых мегаспорофиллами или семенными чешуя ми. Эти чешуи собраны в шишки. У покрытосеменных семязачатки, а следовательно, и семена, заключены в особые структуры, т. е. лучше защищены.
2.7.4. Отдел Coniferophyta (хвойные)
Основные признаки Coniferophyta суммированы в табл. 2.10.
Хвойные (голосеменные) — процветающая группа растений, распространенных по всему
земному шару; на их долю приходится пример но одна треть всех лесов планеты. Это деревья и кустарники, в основном вечнозеленые с игловидными листьями. Подавляющее число видов обитают в высоких широтах и распространяются на север дальше всех других деревьев. Хвойные имеют большое хозяйственное значение в качестве «мягкой древесины», используемой не только для получения лесо- и пиломатериалов, но и для получения смолы, скипидара и древесной мезги. К хвойным относятся сосны, лиственницы (с опадающей на зиму листвой), пихты, ели и кедры. Типичный представитель хвойных — сосна обыкновенная (Pinus sylvestris).

Этот вид распространен на всей территории центральной и северной Европы, бывшего СССР и Северной Америки. Он был интродуцирован и в Великобританию, но в природных условиях растет только в Шотландии. Это красивое величественное дерево высотой до 36 м с характерной отслаивающейся корой розовато или желтовато-коричневого цвета выращивают и для декоративных целей, и для получения пило- и лесоматериалов. Сосны чаще всего растут на песчаных или бедных горных почвах, и поэтому корневая система у них обычно расстилается в поверхностных слоях почвы и сильно ветвится. Внешний вид сосны показан на рис. 2.39.
Каждый год из мутовки боковых почек на верхушке ствола вырастает новая мутовка ветвей. Характерный заостренный на конус облик сосны и других хвойных обусловлен тем, что мутовки более коротких (более молодых) веток на верхушке книзу постепенно сменяются все более длинными (более старыми). С возрастом нижние ветви отмирают и отпадают; поэтому крона у старых деревьев обычно сохраняется только на верхушке (рис. 2.39).
Главные ветви и ствол продолжают расти из года в год в результате роста верхушечной почки. Поэтому говорят, что для хвойных характерен неограниченный рост. Чешуевидные листья расположены спирально; в пазухах таких листьев находятся почки, из которых развиваются короткие веточки (длиной 2–3 мм), называемые укороченными побегами. Это — стебли с ограниченным ростом, на верхушке которых растет по два листа. Как только побег вырастает, чешуевидный лист у основания отпадает и на его месте остается рубец. Листья похожи на иголки, что уменьшает площадь их поверхности, а следовательно, и потери воды. Кроме того, листья по крыты толстой восковой кутикулой, а устьица погружены глубоко в ткань листа — еще одна адаптация для сохранения воды. Эти ксероморфные черты позволяют сводить к минимуму потери воды из вечнозеленых листьев во время холодных сезонов, когда вода может замерзнуть, или когда ее трудно извлечь из почвы. Через два-три года укороченные побеги отпадают вместе с листьями и на их месте остается еще один рубец.
Дерево представляет собой спорофитное поколение. Весной на одном и том же дереве образуются мужские и женские шишки. Диаметр мужских шишек составляет около 0,5 см; они округлые и располагаются группами у основания новых побегов под верхушечной почкой.
Развиваются они в пазухах чешуевидных листьев вместо укороченных побегов. Женские шишки появляются в пазухах чешуевидных листьев на конце новых сильных побегов на некотором рас стоянии от мужских шишек и располагаются более беспорядочно. Поскольку полное развитие шишки занимает три года, шишки очень различаются по размерам, и на одном дереве можно обнаружить шишки от 0,5 до 6 см величиной.
Молодые шишки имеют зеленый цвет, но на второй год они становятся коричневыми или красновато-коричневыми. И мужские, и женские шишки состоят из плотно прижатых друг к другу спорофиллов (модифицированных листьев), расположенных спирально вокруг централь ной оси (рис. 2.39).
На нижней поверхности каждого спорофилла мужской шишки находятся два микроспорангия, или пыльцевых мешка. Внутри каждого пыльцевого мешка происходит мейотическое деление, приводящее к образованию гаплоидных пыльцевых зерен, или микроспор. В них находятся мужские гаметы. У пыльцевых зерен имеется по два больших воздушных мешка, облегчающих их распространение ветром. В мае шишки становятся совсем желтыми из-за пыльцы, которая целым облаком вылетает из них. В конце лета они увядают и отпадают.
Каждый спорофилл женской шишки состоит из нижней кроющей чешуйки и более крупной верхней чешуи, несущей семязачатки. На верхней поверхности крупной чешуи бок о бок расположены два семязачатка, внутри которых происходит образование женских гамет. Опыление происходит уже на первом году развития шишки, но оплодотворение задерживается до следующей весны, когда вырастут пыльцевые трубки. Из оплодотворенных семязачатков образуются крылатые семена. Созревание семян продолжается в течение второго года, и лишь на третий год они высыпаются. К этому времени шишка становится относительно крупной и одревесневает, а чешуи отгибаются наружу, обнажая семена перед тем как их развеет ветром.
2.7.5. Отдел Angiospermophytae (покрытосеменные, или цветковые растения)
Основные признаки покрытосеменных перечислены в табл. 2.10.

Покрытосеменные лучше других растений адаптированы к жизни на суше. Появившись в меловом периоде примерно 135 млн. лет назад, они быстро вытеснили хвойные, заняв главенствующее положение среди растительности плане ты и освоив самые разнообразные местообитания. Некоторые из покрытосеменных вернулись к пресноводному, а несколько видов — солоновато-водному образу жизни.
Одна из самых характерных особенностей покрытосеменных, помимо закрытых семян, о которых уже говорилось, это — появление цветков вместо шишек. Наличие цветков позволило этим растениям привлечь для опыления насекомых, а иногда даже птиц и летучих мышей. Яркая окраска цветков, аромат, съедобная пыльца и нектар — все это средства для привлечения животных-опылителей. В некоторых случаях эволюция опылителей и цветков шла параллель но, что привело к возникновению многих высокоспециализированных и взаимовыгодных отношений. Адаптации цветка, как правило, были направлены на максимальное увеличение шансов для переноса пыльцы насекомыми, и поэтому этот процесс более надежен, чем опыление ветром. Растениям, опыляемым насекомыми, не нужны такие большие количества пыльцы, как при опылении ветром. Тем не менее многие цветковые растения приспособились к опылению ветром.
Двудольные и однодольные
Покрытосеменные растения делятся на две большие группы, которым можно дать статус классов. Чаще всего эти две группы называют однодольными и двудольными. В табл. 2.11 перечислены основные признаки, по которым они различаются (см. также рис. 2.40). Согласно современным представлениям, однодольные, вероятно, произошли от двудольных.

Покрытосеменные бывают травянистые (т. е. не одревесневшие) и деревянистые. К последним относятся кустарники и деревья. У этих растений образуется большое количество вторичной ксилемы (древесины), которая выполняет опорную функцию и, кроме того, служит проводящей тканью. Ксилема возникает в результате активности проводящих пучков камбия. Этот камбий представляет собой слой клеток, расположенных между ксилемой и флоэмой в стеблях и корнях. Клетки камбия сохраняют способность к делению. Образующаяся при этом новая ксилема называется вторичной ксилемой или древесиной.
Травянистые растения, или травы, полагаются для опоры только на тургесцентность клеток и на большое количество механических тканей, таких как колленхима, склеренхима и ксилема; неудивительно поэтому, что и сами они не очень велики. У травянистых растений либо совсем нет камбия, либо, если он и имеется, его активность незначительна.
Многие травянистые растения однолетние, т. е. весь цикл развития от семени до семени у них завершается за один год. У некоторых травянистых растений образуются многолетние органы типа луковиц, клубнелуковиц или клубней, благодаря которым растение перезимовывает или же переживает периоды неблагоприятных условий, например засуху.
Такие травянистые растения могут быть двулетними или многолетними. В первом случае они и образуют семена, и отмирают на второй год, во втором — живут много лет. Кустарники и деревья — растения многолетние. Они могут быть либо вечнозелеными, либо листопадными. Вечно зеленые растения образуют и сбрасывают листву круглый год, поэтому на растениях всегда есть листья. Листопадные полностью сбрасывают листья в холодное или засушливое время года.
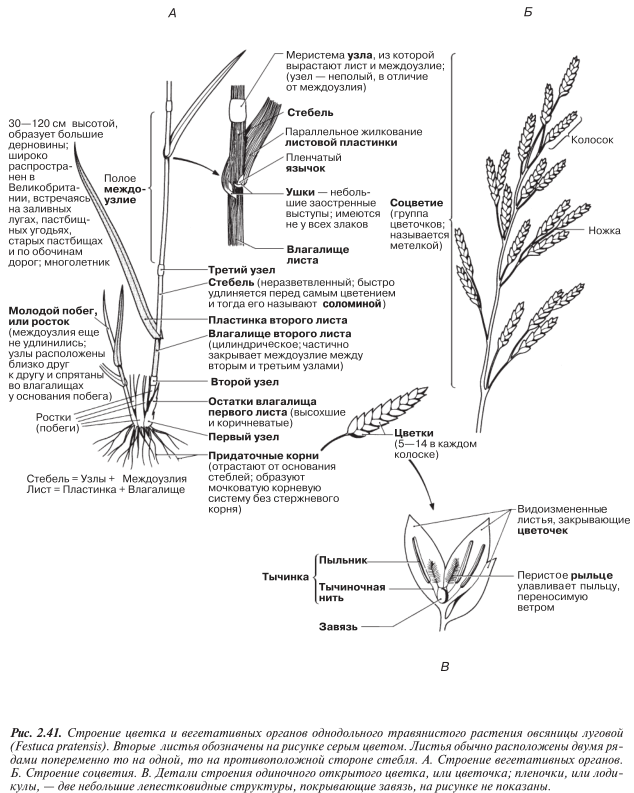
Разнообразие покрытосеменных проиллюстрировано на рис. 2.41–2.44 на примере типичных представителей этой группы растений.



2.7.6. Адаптации растений к жизни на суше
Теперь, когда мы познакомились с отличительными признаками четырех основных групп растений, а именно моховидных, папоротниковидных, голосеменных и покрытосеменных (цветковых), нам легче представить эволюционный прогресс, сделанный растениями в процессе адаптации к жизни на суше.
Проблемы адаптации
Пожалуй, наитруднейшей проблемой, которую надо было как-то преодолеть, чтобы перейти от водного образа жизни к наземному, была проблема обезвоживания. Любое растение, незащищенное тем или иным способом, например не покрытое восковой кутикулой, очень скоро вы сохнет и несомненно погибнет. Даже если преодолеть эту трудность, останутся другие нерешенные проблемы. И прежде всего вопрос о том, как успешно осуществить половое размножение.
У первых растений в размножении участвовали мужские гаметы, способные приблизиться к женским гаметам, только плавая в воде.
Обычно считают, что первые растения, освоившие сушу, произошли от зеленых водорослей, у отдельных из наиболее эволюционно продвинутых представителей которых появились ре продуктивные органы, а именно архегонии (женские) и антеридии (мужские); в этих органах были спрятаны, а, следовательно, и защищены гаметы. Это обстоятельство и ряд других вполне определенных приспособлений, помогающих избежать высыхания, позволили некоторым представителям зеленых водорослей завладеть сушей.
Одна из важнейших эволюционных тенденций у растений — это постепенно увеличивающаяся независимость их от воды.
Ниже перечислены основные трудности, которые связаны с переходом от водного к на земному существованию:
- 1. Обезвоживание. Воздух — это среда, способствующая высыханию, а вода необходима для жизни по целому ряду причин. Следовательно, возникает необходимость в приспособлениях для получения и запасания воды.
- 2. Размножение. Нежные половые клетки должны быть защищены, а подвижные мужские гаметы (спермии) могут встретиться с женскими гаметами только в воде.
- 3. Опора. В отличие от воды воздух не может служить опорой растениям.
- 4. Питание. Растениям необходимы свет и диоксид углерода (CO2) для фотосинтеза, поэтому хотя бы часть растения должна возвышаться над землей. Однако минеральные соли и вода находятся в почве или на ее поверхности, и, чтобы эффективно использовать эти вещества, часть растения должна находиться в земле и расти в темноте.
- 5. Газообмен. Для фотосинтеза и дыхания нужно, чтобы обмен диоксида углерода и кислорода происходил не с окружающим раствором, а с атмосферой.
- 6. Факторы окружающей среды. Вода, особенно, когда ее так много, как, скажем, в озере или в океане, обеспечивает высокое постоянство условий окружающей среды.
Наземная же среда обитания в гораздо большей степени характеризуется изменчивостью таких важных факторов, как температура, интенсивность освещения, концентрация ионов и pH.
Печеночники и мхи
Мхи хорошо приспособились к распространению спор в наземных условиях: оно зависит от высыхания коробочки и рассеивания мелких легких спор ветром. Однако эти растения все еще находятся в зависимости от воды по следующим причинам:
- 1. Вода необходима им для размножения, поскольку спермии должны подплывать к архегониям. У этих растений возникли адаптации, позволяющие им высвобождать спермии только во влажной среде, потому что только в такой среде вскрываются антеридии. Эти растения частично приспособились к наземной жизни, поскольку га меты у них образуются в защитных структурах — антеридиях и архегониях.
- 2. У них нет специальных опорных тканей, и поэтому рост растения вверх ограничен.
- 3. У моховидных нет корней, способных далеко проникать в субстрат, и они могут жить только там, где на поверхности почвы или в ее верхних слоях имеется достаточно влаги и минеральных солей. Однако у них имеются ризоиды, которыми они прикрепляются к грунту; это — одна из адаптаций к жизни на твердом субстрате.
Семенные растения — хвойные и цветковые
Одна из основных трудностей, с которой сталкиваются растения на суше, связана с уязвимостью гаметофитного поколения. Например, у папоротников гаметофит — это нежный заросток, который образует мужские гаметы (спермии), нуждающиеся в воде, чтобы достичь яйцеклетки.
Однако у семенных растений гаметофит защищен и сильно редуцирован.
Семенные растения обладают тремя важны ми преимуществами: во-первых, разноспоровостью; во-вторых, появлением неплавающих мужских гамет и, в-третьих, образованием семян.
РАЗНОСПОРОВОСТЬ И НЕПЛАВАЮЩИЕ МУЖСКИЕ ГАМЕТЫ.
Очень важную роль в эволюции растений сыграло возникновение некоторых папоротников и их близких родичей, образующих споры двух типов.
Явление это называют разноспоровостью, а растения — разноспоровыми. Все семенные растения относятся к разноспоровым. Они образуют крупные споры, называемые мегаспорами, в спорангиях одного типа (мегаспорангиях) и мелкие споры, называемые микроспорами, — в спорангиях другого типа (микроспорангиях). Прорастая, споры образуют гаметофиты (рис. 2.34). Мегаспоры развиваются в женские гаметофиты, микроспоры — в мужские. У семенных растений гаметофиты, образуемые мегаспорами и микро спорами, очень малы по размерам и никогда не высвобождаются из спор. Таким образом, гаметофиты оказываются защищенными от высыхания, что представляет собой важное эволюционное достижение. Однако спермии из мужского гаметофита все еще должны перемещаться к женскому гаметофиту, что значительно облегчается рассеиванием микроспор. Будучи очень мелкими, они могут образовываться в больших количествах и разноситься ветром далеко от родительского спорофита. Случайно они могут оказаться в тесной близости от мегаспоры, которая у семенных растений не отделяется от родительского спорофита (рис. 2.45).
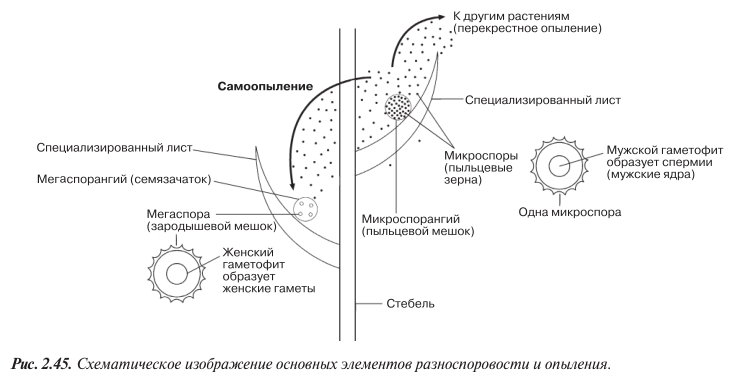
Именно таким путем и происходит опыление у растений, пыльцевые зерна которых представляют собой микроспоры. В пыльцевых зернах образуются мужские гаметы.
У семенных растений возникло еще одно эволюционное преимущество. Мужским гаметам не нужно больше подплывать к женским гаметам, поскольку у семенных растений появились пыльцевые трубки. Они развиваются из пыльцевых зерен и растут в направлении женских гамет. По этой трубке мужские гаметы достигают женской гаметы и оплодотворяют ее. Плавающие спермии больше не образуются, в оплодотворении участвуют только мужские ядра. Следовательно, у растений выработался механизм оплодотворения, независимый от воды.
Это и послужило одной из причин, по которой семенные растения столь превзошли другие растения в освоении суши. Первоначально опыление происходило только с помощью ветра — процесс довольно случайный, сопровождающийся большими потерями пыльцы. Однако уже на ранних этапах эволюции примерно 300 млн. лет назад в каменноугольном периоде, появились летающие насекомые, а с ними и возможность более эффективного опыления. Цветковые растения широко используют опыление насекомыми, тогда как у хвойных все еще преобладает опыление ветром.
СЕМЕНА.
У ранних разноспоровых растений мегаспоры высвобождались из родительского спорофита подобно микроспорам. У семенных же растений мегаспоры не отделяются от родительского растения, оставаясь в мегаспорангиях, или семязачатках (рис. 2.45). Семязачаток содержит женскую гамету. После оплодотворения женской гаметы семязачаток называют уже семенем. Таким образом, семя — это оплодотворенный семязачаток. Наличие семязачатка и семени дает определенные преимущества семенным растениям.
- 1. Женский гаметофит защищен семязачатком. Он полностью зависит от родительского спорофита и в отличие от свободно живущего гаметофита нечувствителен к обезвоживанию.
- 2. После оплодотворения в семени образуется запас питательных веществ, получаемых гаметофитом от родительского спорофитного растения, от которого он по-прежнему не отделен. Этот запас используется развивающейся зиготой (следующим спорофитным поколением) после прорастания семени.
- 3. Семена предназначены для того, чтобы переживать неблагоприятные условия, и остаются в состоянии покоя до тех пор, пока условия не станут благоприятными для прорастания.
- 4. У семян могут развиваться различные приспособления, облегчающие их распространение.
Семя представляет собой сложную структуру, в которой собраны клетки трех поколений — родительского спорофита, женского гаметофита и зародыша следующего спорофитного поколения. Родительский спорофит дает семени все, что нужно для жизни, и только после того, как семя полностью созреет, т. е. накопит запас питательных веществ для зародыша спорофита, оно отделяется от родительского спорофита.
2.7.7. Краткое перечисление адаптаций семенных растений к жизни на суше
Основные преимущества семенных растений над всеми остальными сводятся к следующему.
- 1. Гаметофитное поколение сильно редуцировано и полностью зависит от хорошо приспособленного к жизни на суше спорофита, внутри которого гаметофит всегда защищен. У других растений гаметофит очень легко высыхает.
- 2. Оплодотворение происходит независимо от воды. Мужские гаметы неподвижны и разносятся внутри пыльцевых зерен ветром или насекомыми. Окончательный перенос мужских гамет к женским происходит с помощью пыльцевой трубки.
- 3. Оплодотворенные семязачатки (семена) остаются некоторое время на родительском спорофите, от которого они получают защиту и пищу прежде, чем будут развеяны.
- 4. У многих семенных растений наблюдается вторичный рост с отложением больших количеств древесины, несущей опорную функцию. Такие растения вырастают в деревья и кустарники, способные эффективно конкурировать в борьбе за свет и другие ресурсы.
Некоторые из важнейших эволюционных тенденций приводятся в обобщенном виде на рис. 2.33. У семенных растений имеются и другие признаки, присущие растениям не только этой группы, но также выполняющие роль адаптаций к жизни на суше.
- 1. Настоящие корни обеспечивают извлечение влаги из почвы.
- 2. Растения защищены от высыхания эпидермисом с водонепроницаемой кутикулой (или пробкой, образующейся после вторичного роста).
- 3. Эпидермис наземных частей растения, особенно листьев, пронизан множеством мельчайших щелей, называемых устьицами, через которые осуществляется газообмен между растением и атмосферой.
- 4. У растений имеются и специализированные адаптации к жизни в жарких засушливых условиях.