Репродуктивные органы цветковых растений
Ботаника. Учебное пособие по специальности «Фармация»
- Морфология цветка. Репродуктивные процессы в цветке
- Соцветия
- Плоды. Морфология и классификация плодов
- Семя
- Распространение плодов и семян
Покрытосеменные, или цветковые растения являются наиболее крупным отделом растительного царства, насчитывающим более 350 семейств, 13 тысяч родов и до 240 тысяч видов. В настоящее время покрытосеменные распространены по всему Земному шару – от тропиков до приполярных пустынь. Автотрофные цветковые растения – важнейший компонент биосферы, от которого зависят биосферные процессы обмена веществ и трансформации энергии, газовый состав атмосферы, климат, водный режим суши, характер процессов почвообразования. На суше цветковые часто доминируют в растительном покрове, только в немногих сообществах и типах растительности они уступают господство по общей массе другим группам растений – в хвойных лесах голосеменным, в некоторых типах тундр и на сфагновых болотах – мохообразным.
Покрытосеменные определяют самую возможность существования большинства наземных животных. Они формируют среду обитания для разнообразных консументов – потребителей первичного органического вещества. Различные группы гетеротрофов связаны с ними трофическими, консортивными связями. В процессе эволюции многие группы гетеротрофов смогли возникнуть только тогда, когда Землю завоевали покрытосеменные и образуемые ими сообщества. Для многих членистоногих и позвоночных характерна сопряженная с цветковыми эволюция. Человек как биологический вид тоже возник и эволюционировал после появления покрытосеменных, отличающихся высоким уровнем продуктивности и биохимическим разнообразием.
Покрытосеменные растения отличаются необычайным полиморфизмом, эволюционной пластичностью и могут существовать в разнообразных условиях обитания. Это единственная группа среди высших растений, представители которой сумели вторично освоить морскую среду, где стали доминировать на огромных акваториях.
Разнообразие размеров и жизненных форм цветковых растений, их приспособлений к конкретным условиям обитания велико. Представитель семейства рясковых – ряска малая (Lemna minor) не превышает 3-5 мм в длину и похожа на свободноплавающие слоевищные растения – мхи или водоросли (рис. 13.1).

Рис. 13.1. Ряска малая (Lemna minor) – миниатюрное цветковое растение.
С другой стороны, гигантские эвкалипты (виды рода Eucalyptus из семейства миртовые Myrtaceae) вырастают высотой до 100 м, длина побегов тропических лиан – ротанговых пальм измеряется сотнями метров. Среди покрытосеменных преобладают автотрофы, но встречаются полупаразиты, паразиты и сапротрофы, полностью утратившие хлорофилл и способность к фотосинтезу. Среди покрытосеменных имеются также эпифиты и эпифиллы (эпифиты, живущие на листьях других растений), деревья, кустарники, кустарнички, полукустарники и травы (многолетники и однолетники), плотоядные растения с их разнообразными приспособлениями для улавливания и переваривания мелких животных (рис. 13.2).

Рис. 13.2. Насекомоядное цветковое растение росянка круглолистная (Drosera rotundifolia).
Отмечены разнообразные приспособления видов цветковых растений к опылению строго определенными агентами, к распространению плодов и семян, к перенесению неблагоприятных климатических периодов. Эти особенности дают цветковым растениям возможность полностью реализовать свой эволюционно-адаптационный потенциал. Ни одна другая группа растений не смогла выработать такого разнообразия приспособлений к определенным факторам среды.
Существенным признаком покрытосеменных считают цветок – отсюда одно из названий отдела – Anthophyta (antos – цветок, phyton – растение). Происхождение цветка как особого органа – это одна из ключевых проблем морфологии и филогении покрытосеменных. Дискуссии по этому вопросу, начавшиеся еще в XVIII в., продолжаются и в настоящее время.
Типичный цветок определяют как видоизмененный побег, обеспечивающий функции спороношения, развития гаметофитов, половое размножение, образование семян в плоде. В истории ботаники отмечено, что в поисках архетипа цветка за первичный примитивный цветок различные ученые принимали разные структуры.
Сторонники эвантовой теории считают первичным крупный обоеполый цветок, опыляемый насекомыми, с большим и часто неопределенным числом свободных частей, и тогда цветки однополые, олигомерные, голые рассматриваются как результат более поздних этапов редукционной эволюции.
Сторонники псевдантовой теории полагают, что эволюционные трансформации выражались в постепенном усложнении структуры цветка от однополых, анемофильных, с небольшим числом частей к обоеполым, полимерным, энтомофильным, с хорошо развитым околоцветником.
Следует учитывать, что в процессе эволюционного возникновения покрытосеменные сначала приобрели комплекс важнейших общих признаков (в первую очередь – уникальные особенности жизненного цикла), а уже потом выработали разные цветки. При этом в каждой эволюционной ветви покрытосеменных были осуществлены различные морфогенетические решения.
Существенными частями цветка являются андроцей, состоящий из одной, нескольких или многих тычинок (микроспорофиллов) и гинецей, который состоит из одного, нескольких или многих плодолистиков, или карпелл (мегаспорофиллов).
Плодолистики (карпеллы) могут быть свободными, и тогда каждый из них, срастаясь краями, образует пестик (один или несколько пестиков в цветке – по числу плодолистиков). При срастании нескольких плодолистиков формируется ценокарпный гинецей, пестик в нем всегда один.
Важнейшая часть пестика – завязь. Наличие завязи (замкнутой полости), в которой развиваются семязачатки (в отличие от голосеменных, у которых семязачатки расположены на поверхности открытых мегаспорофиллов), обычно считают важнейшим признаком покрытосеменных, откуда и другое название отдела – Angiospermae (от греческого angeion – сосуд, вместилище, sperma – семя). Полагают, что завязь обеспечивает лучшую защиту семяпочек (семязачатков). В полости завязи семязачатки оказываются в условиях влажной камеры, оптимальных для их развития. Благодаря образованию завязи расширяются и обогащаются возможности распространения растений: из цветка формируется плод, при диссеминации расселяются не только отдельные семена, но и целые плоды, у которых возникают разнообразные приспособления к распространению широким кругом агентов.
У цветковых растений и некоторых голосеменных спермин (мужские неподвижные половые клетки) доставляются к яйцеклетке с помощью особой клетки – пыльцевой трубки, которая обеспечивает высокую точность попадания спермия в зародышевый мешок (то есть к женскому гаметофиту с яйцеклеткой). Семязачатки у покрытосеменных растений скрыты в завязи пестика, поэтому необходимо приспособление для улавливания пыльцы. Таким аппаратом у покрытосеменных служит рыльце на верхушке столбика пестика (стилодия). Наличие рыльца – важная особенность покрытосеменных растений.
Архегониальные растения (папоротникообразные, голосеменные) характеризуются тенденцией к возникновению разноспоровости, которая сопровождается редукцией полового поколения в жизненном цикле. Покрытосеменные, у которых редуцированы гаметангии, представляют собой завершающий этап на этом пути. Предельное сокращение процессов микро- и мегаспорогенеза и ускорение развития гаметофитов, связанное с их редукцией, были, вероятно, важными эволюционными факторами. Женский гаметофит цветковых растений представлен зародышевым мешком, состоящим всего из 7 клеток, а мужской гаметофит представлен пыльцевым зерном (пылинкой), в которой только 2 клетки – сифоногенная и спермагенная (дающая 2 спермия).
Уникальной особенностью покрытосеменных является двойное оплодотворение, в котором участвуют 4 клетки: два спермия, яйцеклетка и центральная клетка зародышевого мешка. В ходе этого процесса один из cnepмиев оплодотворяет яйцеклетку, образуя диплоидную зиготу, которая впоследствии развивается в зародыш семени. Второй спермий сливается с диплоидным ядром центральной клетки зародышевого мешка, образуя триплоидную зиготу. Эта зигота впоследствии дает начало триплоидному эндосперму семени. Общий для всех покрытосеменных уникальный процесс двойного оплодотворения указывает на их эволюционное единство и монофилетическое становление.
Одно из следствий двойного оплодотворения – формирование питательной ткани – полиплоидного эндосперма (в отличие от гаплоидного эндосперма голосеменных, представляющего остаток женского гаметофита). Биологическое значение полиплоидного состояния запасающей ткани связано с тем, что оно обеспечивает более благоприятные условия для развития зародыша и поэтому могло иметь важную эволюционную роль.
На тканевом уровне подавляющее большинство покрытосеменных характеризуется наличием в ксилеме сосудов, тогда как почти у всех голосеменных проводящие элементы древесины представлены только трахеидами. Только у немногих цветковых проводящая система ксилемы гомоксильная, бессосудистая, унаследовавшая примитивность от предковых голосеменных форм.
Паренхиматизация цветковых позволила выработать специальные запасающие ткани и травянистые жизненные формы. В отличие от древесных растений, у трав возможно сокращение онтогенеза и ускорение темпов эволюции; травянистые растения способны существовать в таких крайних условиях среды, в которых деревья и кустарники жить не могут.
Остается нерешенной проблема геологического возраста цветковых растений. Считалось, что покрытосеменные растения возникли раньше мелового периода – в триасе или палеозое (255-210 млн. лет назад). Предполагалось, что первоначально небольшая, эта группа существовала в таких условиях, которые не благоприятствовали фоссилизации их остатков, например, в горных районах или в пустынях. Позже, в меловой период (145-63 млн. лет назад), покрытосеменные дали мощную вспышку развития и сформировали свое современное разнообразие.
Этому взгляду противоречат объективные данные палеоботаники. Многочисленные указания на нахождение ископаемых остатков покрытосеменных в домеловых осадочных породах в настоящее время достоверно не доказаны. Имеются косвенные указания на возможность существования покрытосеменных в юре (210-145 млн. лет назад). Например, с этого времени известны некоторые формы насекомых, современные родственники которых настолько тесно связаны с цветковыми растениями, что допустить их независимое от последних существование трудно.
Древнейшие, достоверно определяемые ископаемые остатки цветковых растений в виде фоссилизованных листьев, цветков, плодов, древесины и пыльцевых зерен встречаются лишь с начала мелового периода (135 млн. лет назад). В этих отложениях они очень немногочисленны и фрагментарны. Еще недавно считалось, что к середине мелового периода (120 млн. лет назад) покрытосеменные появились в массе и выступали как «победители в борьбе за существование» с голосеменными и папоротникообразными. Сейчас палеоботаники отодвигают начало расцвета покрытосеменных на более позднее время – конец мелового периода (80-63 млн. лет назад).
Предполагают, что родиной цветковых могли быть тропические или субтропические регионы Земли. Исследователи считают, что это были районы тропической Юго-Восточной и Восточной Азии и Меланезия. В этом районе сосредоточены древние и считающиеся примитивными таксоны цветковых. Вероятно, что отсюда шла миграция древнейших цветковых в другие области Земли. Еще одна точка зрения заключается в признании центром происхождения покрытосеменны[ древнего континента Южного полушария – Гондваны. Считается, что цветковые могли возникнуть и расселиться по этому обширному континенту до начала раскола Гондваны на современные континенты Южного полушария.
К началу XX в. сложились две противостоящие друг другу системы взглядов на происхождение покрытосеменных от более примитивных предковых форм растений. Одна из них, связанная с именами австрийского ботаника Р. Веттштейна и классика немецкой ботаники А. Энглера, была основана на представлении о происхождении цветковых от эфедроподобных или гнетоподобных голосеменных предков. Р.Веттштейном была разработана оригинальная концепция происхождения цветка – так называемая псевдантовая гипотеза (от греческого «псевдантиум» – ложный цветок). Согласно этой гипотезе цветок представляет собой собрание мелких разнополых стробилов, претерпевших в процессе эволюции ряд преобразований: редукцию, сближение и срастание. На основе этой гипотезы наиболее архаичными следует считать семейства с тычиночными и пестичными невзрачными ветроопыляемыми цветками (ивовые, казуариновые).
Другая концепция восходит к построениям И.В.Гёте и А.П.Декандоля, то есть к концу XVIII – началу XIX веков. В эпоху филогенетической систематики это направление ознаменовалось появлением стробилярной (от греческого «стробилюс» – шишка хвойного)гипотезы, которую сформулировали Е.Арбер и Дж. Паркин. Другое название этой гипотезы – эвантиевая (от греческого «эвантиум» – настоящий цветок). Согласно стробилярной гипотезе наиболее вероятными предками покрытосеменных были мезозойские голосеменные растения – беннеттиты. С этих позиций цветок – это метаморфизированный (видоизмененный) укороченный спороносный побег, первоначально напоминавший шишку голосеменных. Мегаспорофиллы, несущие семязачатки, в процессе метаморфоза превратились в плодолистики (карпеллы), а несущие пыльники микроспорофиллы – в тычинки. Эти видоизменения многие исследователи связывают с приспособлением покрытосеменных к насекомоопылению (энтомофилии). Примитивные цветки имели значительные размеры и выпуклое цветоложе, на котором располагались многочисленные микро- и мегаспорофиллы. Микроспорофиллы эволюционировали в тычинки, а мегаспорофиллы – в карпеллы (плодолистики), которые образовали пестики. Цветки приобрели строение, сходное с тем, которое наблюдается у современных магнолиевых, лютиковых, кувшинковых. Поэтому перечисленные семейства обычно располагаются в основании системы покрытосеменных как наиболее древние и архаичные. Дальнейшая эволюция цветка в пределах покрытосеменных имела редукционный характер.
Обе эти гипотезы являются фолиарными (от латинского «фолиум» – лист), так как исходят из представлений, что цветки образовались из листостебельных спороносных побегов. Фолиарным гипотезам противопоставляются различные теломные (от греческого «телома» – конечные побеги) гипотезы (например, теория гонофилла Р. Мелвилла), широко распространенные в 60-70-е годы ХХ в. Согласно этим гипотезам, все части цветка могут быть выведены из теломов – осевых структур, свойственных ископаемым растения – риниевым.
Палеоботаника в последние десятилетия дала много нового материала о древнейших, ранне- и среднемеловых покрытосеменных. Эти данные позволяют сделать несколько заключений. Во-первых, уже самые ранние из известных нам цветковых отличаются большим полиморфизмом. Очень разнообразны пыльцевые зерна по форме, размерам, скульптуре поверхности, характеру поровости, числу и строению апертур. Они обнаруживают сходство с пыльцевыми зернами порядков и семейств современных растений. Листья меловых цветковых также весьма разнообразны и напоминают современные фикусы, протейные, дубы, акации, аралию, подорожник, известны и специализированные щитовидные листья.
Большим полиморфизмом отличаются и органы размножения – цветки и плоды. У меловых цветковых известны как одиночные и относительно крупные цветки, так и мелкие, собранные в разнообразно устроенные соцветия с простым или двойным околоцветником, с апокарпным или ценокарпным гинецеем, с верхней или нижней завязью, с разным числом тычинок и плодолистиков. Наряду со вскрывающимися плодами типа листовок обнаружены и не-вскрывающиеся плоды. Вместе с древесными формами в меловую эпоху существовали многочисленные травянистые покрытосеменные.
Во-вторых, если раньше часто утверждали, будто в меловую эпоху покрытосеменные были представлены современными таксонами, то сейчас большинство исследователей считают, что раннемеловые Angiospermae принадлежали к вымершим порядкам и семействам. Этот факт важен как доказательство того, покрытосеменные не оставались неизменными с момента зарождения.
В целом сейчас большинство специалистов считают, что предков Angiospermae нужно искать среди каких-то древних и малоспециализированных голосеменных растений. Не оставлена идея их генетической связи с беннеттитовыми. Допускают, что магнолиевые (как наиболее примитивные цветковые) лишь сохранили наибольшее число архаичных признаков, но сами, как и все прочие таксоны цветковых, произошли от какой-то вымершей, еще более древней гипотетической группы покрытосеменных.
В настоящем учебном пособии принята система цветковых растений, разработанная А.Л.Тахтаджяном в 1987 г. В данной системе древнейшей группой покрытосеменных считается порядок магнолиевых (Magnoliales), от предков которых, вероятно, произошли современные покрытосеменные. В этом смысле приведенная система монофилетична (рис. 13.3).
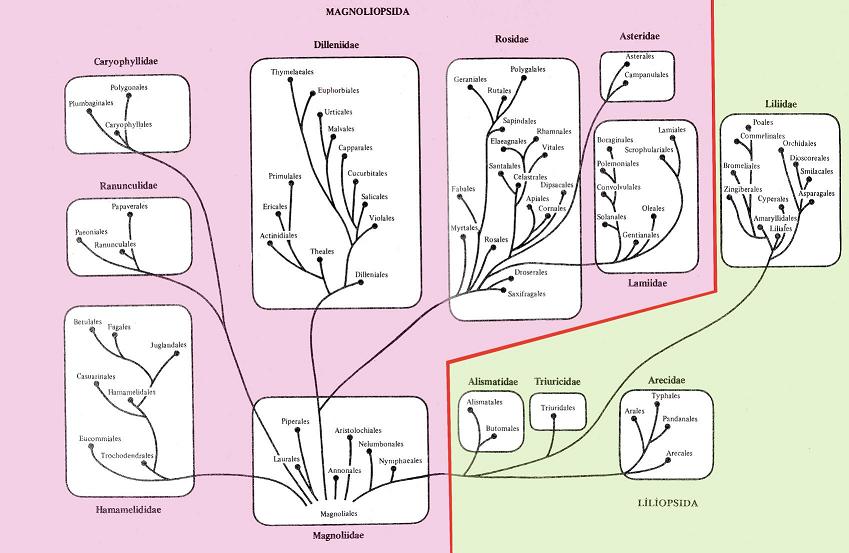
Рис. 13.3. Система покрытосеменных растений (по А.Л.Тахтаджяну).
Цветок – особый побег покрытосеменных растений, ответственный за формирование спор двух типов (микро- и мегаспор), развитие мужского и женского гаметофитов, образование гамет (яйцеклеток и спермиев), опыление, оплодотворение и формирование семян. Цветок в процессе своего развития превращается в плод с семенами. Во многих отношениях он уникален и столь характерен для этой группы, что покрытосеменные нередко называют цветковыми растениями. Исключительная роль цветка как особой морфологической структуры связана с тем, что в нем полностью совмещены все процессы бесполого и полового размножения. В цветках осуществляются микро- и мегаспорогенез, развитие гаметофитов, микро- и мегагаметогенез, опыление, оплодотворение и формирование зародышей нового поколения спорофитов. Завершается онтогенез цветка образованием плода с семенами. Особенности строения цветка обеспечивают осуществление всех этих функций с наименьшими затратами пластических веществ и энергии.
13.5. Семя
Семя – особая структура семенных растений (голосеменных и покрытосеменных), развивающаяся из семязачатка после процесса двойного оплодотворения, реже без оплодотворения (апомиксис), обеспечивающая расселение потомства. При формировании семя заключено в перикарпий и является частью плода. В отличие от споры, обеспечивающей расселение споровых растений, семя обладает рядом преимуществ, возникших в результате прогрессивной эволюции. Семя представляет собой многоклеточную структуру, объединяющую зачаточное растение (зародыш), запасающую ткань и защитный покров. Этим семя существенно отличается от споры, где вещества, необходимые для развития будущего растения-гаметофита, содержатся в единственной микроскопической клетке. Физиологически спора и семя также существенно различаются. Спора прорастает немедленно при поступлении в клетку влаги. Многие семена имеют различной длительности период покоя, в течение которого они не способны к активной жизнедеятельности и образованию проростка. Иными словами, семена как единицы расселения растений во всех отношениях значительно более надежны и универсальны, чем споры. Образование из семязачатка семени начинается с того, что зигота, располагающаяся в семязачатке, вытягивается в длину и делится поперек перегородкой. Одна из клеток образует так называемый подвесок, или суспензор, другая – собственно зародыш. Подвесок содействует питанию зародыша, погружая его в эндосперм, а нередко приобретает свойства гаустории – присоски. Вторая клетка многократно митотически делится и в конечном итоге образует сформировавшийся зародыш. Начало эндосперму дает триплоидное ядро, образовавшееся в результате слияния диплоидного вторичного ядра зародышевого мешка и одного из спермиев. Деление этого ядра дает всю массу питательной ткани – эндосперма. Степень развития эндосперма у разных таксонов неодинакова. Как правило, чем примитивнее в эволюционном отношении систематическая группа, тем лучше развит у нее эндосперм. Редукция эндосперма обычно связана с увеличением относительных размеров зародыша. С увеличением его размеров запасные вещества обычно накапливаются в самом зародыше. В процессе развития женского гаметофита, а затем зародыша и эндосперма мегаспорангий (нуцеллус семязачатка), обычно разрушается, а его запасные вещества используются. Однако у некоторых групп покрытосеменных эта ткань частично сохраняется, превращаясь в диплоидную запасающую ткань, физиологически аналогичную эндосперму. Эта ткань носит название перисперма (от греческого «пери» – около, возле) и отмечена для семян представителей семейств перцевых, гвоздичных.
Зрелые семена различаются по форме, величине, структуре поверхности, окраске и по внутреннему строению. Очень разнообразна форма семян – шаровидная, дисковидная, эллипсоидная, линейная. Обычно семена невелики по размерам, лишь изредка достигая в длину немногих сантиметров. Очень мелкие семена, почти неразличимые простым глазом, свойственны видам семейства орхидных и многим паразитическим растениям. Крупные семена встречаются у многих бобовых, конского каштана, некоторых тыквенных и ряда пальм. Семена тропического бобового моры маслоносной (Mora oleifera) достигают 16 см в поперечнике, у сейшельской пальмы (Lodoicea maldivica) – 50 см в диаметре и весят до 20 кг.
Семена цератонии стручковой (Ceratonia siliqua) из семейства бобовых очень постоянны по массе и служили у ювелиров мерой веса (1 карат) для драгоценных камней. Поверхность семян может быть гладкой, блестящей, но часто бывает шероховатой, бороздчатой, ребристой, ямчатой, опушенной волосками. Различают семена и по окраске. Особенно разнообразны по окраске семена бобовых. Семена часто снабжены разного рода придатками (присемянниками), выростами (например, крыловидными), иногда пучками волосков. Обычно семена прикреплены в плоде к фуникулусу, но изредка – непосредственно к плаценте. Общий план строения семени определяется типом семязачатка, из которого оно возникло. Основные структурные части зрелого семени: семенная кожура, питательная (запасающая) ткань и зародыш (рис. 13.21).
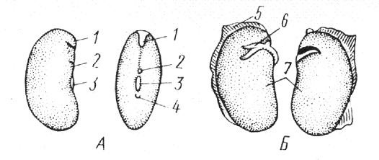
Рис. 13.21. Семя фасоли (Phaseolus vulgaris). А – общий вид; Б – зародыш: 1 – след халазы, 2 – след микропиле, 3 – рубчик, 4 – семенной шов, 5 – семенная кожура, 6 – почечка, 7 – семядоля.
Семенная кожура, или спермодерма, формируется главным образом за счет интегументов семязачатка, реже – за счет разрастания тканей халазы. У большинства растений семенная кожура плотно окружает семя и служит основным защитным покровом, препятствующим его иссушению и преждевременному насыщению влагой. Структурные особенности семенной кожуры связаны со способами распространения и прорастания семян. Они имеют большое значение для систематики. У семян, развивающихся во вскрывающихся плодах, в семенной кожуре часто образуется защитный слой из склерифицированных клеток. Иногда наружный слой кожуры становится мясистым и сочным, что привлекает птиц и млекопитающих и способствует распространению семян. На поверхности семени заметен рубчик – след, остающийся на месте прикрепления семязачатка к фуникулюсу и внутренней поверхности завязи. Морфологические особенности рубчика – форма, размеры, окраска имеют большое значение в систематике растений, а также широко используются в семеноведении при характеристике и определении семян.
Многим семенам цветковых растений свойственно особое образование, имеющее вид мясистых наростов, пленок или бахромы. Оно развивается в различных частях семени и получило название присемянника, или ариллуса (от латинского «ариллус» – сушеный виноград). Природа присемянника различна. Иногда он возникает в результате разрастания тканей фуникулуса, обрастает семя частично или полностью, плотно прилегая к семенной кожуре, но не срастается с нею.
В иных случаях ариллус – производное наружного интегумента семязачатка. Присемянники, располагающиеся близ микропилярного следа семени, известны под названием карункулы. Присемянники большей частью ярко окрашены, играют важную роль в распространении семян и тем самым – в расселении растений. Канал, или углубление в семенной кожуре, являющееся остатком микропиле семязачатка, называется микропилярным следом. Остаток халазы на противоположном конце семени именуется халазальным следом. Через микропилярный след при прорастании семени выходит корешок. Помимо рубчика, микропилярного и халазального следов на семенной кожуре обычно можно заметить особое утолщение, называемое ребром семени или его швом. Шов возникает в той части фуникулуса, которая у некоторых типов семязачатков сливается с интегументом.
Питательной тканью в семенах может быть эндосперм и перисперм. У 85% видов в семенах встречается эндосперм (семена с эндоспермом), реже – перисперм (семена с периспермом), еще реже – обе питательные ткани одновременно (семена с эндоспермом и периспермом). У некоторых таксонов специальные питательные ткани полностью отсутствуют и тогда запасные вещества откладываются непосредственно в зародыше (семена без эндосперма). Консистенция питательной ткани различна: твердая, жидкая, слизистая. Твердый, но снабженный глубокими складками и бороздами эндосперм называется руминированным. Чаще всего в питательной ткани накапливаются углеводы в виде зерен вторичного крахмала, реже липиды в виде капелек жирного масла. В семенах всегда имеются также белки, что особенно важно при прорастании, и фосфорное соединение фитин, которому приписывают роль стимулятора в метаболических (обменных) процессах при прорастании.. В зависимости от химического состава преобладающих запасных веществ семена разделяют на крахмалистые (пшеница, кукуруза, рис и другие злаки), масличные (подсолнечник, лен, арахис, соя) и белковые (бобовые). Выделяют несколько типов семян (рис. 13.22).
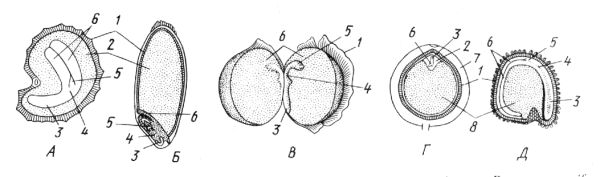
Рис. 13.22. Типы семян. А – с эндоспермом, окружающим зародыш (мак Papaver somniferum, Б – с эндоспермом, примыкающим к зародышу (пшеница Triticum aestivum); В – с запасными веществами, отложенными в семядолях зародыша (горох Pisum sativum); Г – с эндоспермом, окружающим зародыш, и мощным периспермом (перец Piper nigrum); Д – с периспермом (куколь Agrostemma githago): 1 – семенная кожура, 2 – эндосперм, 3 – корешок, 4 – стебелек, 5 – почечка, 6 – семядоли, 7 – околоплодник, 8 – перисперм.
Зародыш (эмбрио) обычно образуется из оплодотворенной яйцеклетки и представляет собой зачаточный спорофит. Процесс формирования зародыша (эмбриогенез) подразделяется на несколько периодов. Семена большинства растений заключают один зародыш. Он чаще всего бесцветен, реже окрашен и тогда содержит хлорофилл. Степень морфологической расчлененности зародыша различна у разных систематических групп. Зародыш в значительной мере составлен из меристемы. Для наиболее примитивных таксонов характерен так называемый недоразвитый зародыш. Он очень мелкий, точечный и формируется при прорастании семени. У эволюционно продвинутых групп зародыш хорошо развит, в его частях могут откладываться питательные вещества, а специальные питательные ткани (эндосперм и перисперм) при этом редуцируются или полностью исчезают. В то же время ряд высокоорганизованных семейств, например, орхидные, имеют зародыши, состоящие из небольшой группы недифференцированных клеток. У большинства цветковых растений ось зародыша состоит из зародышевых корешка и стебелька. У голосеменных и цветковых двудольных растений к верхней части стебелька прикрепляются семядоли – первые листья зародыша. Часть стебелька, располагающаяся ниже семядолей, называется гипокотилем, выше – эпикотилем. Верхушка стебелька завершается почечкой.
В семени растений корешок всегда направлен к следу микропиле. Из него образуется главный корень нового спорофита. У части семян гипокотиль и эпикотиль при прорастании способны удлиняться и выносить семядоли на поверхность (надземное прорастание семян). У двудольных семядолей обычно две, очень редко три или четыре, у однодольных только одна, у голосеменных их чаще всего несколько (от 2 до 15). Семядоли – первые листья зародыша. Считается, что в процессе эволюции цветковых односемядольный зародыш произошел от двусемядольного. При надземном прорастании семядоли зеленеют и способны к фотосинтезу, а при подземном – служат хранилищем питательных веществ (например у лещины, дуба) или выполняют функцию гаустории (структуры, поглощающей питательные вещества). В семенах с эндоспермом они подают питательные вещества в надземную часть проростка. Почечка представляет собой зачаток главного побега растения.
Рост семени заканчивается незадолго до завершения его полного физиологического развития. Несколько позднее прекращается приток питательных веществ и снижается активность растительных гормонов (фитогормонов). По мере того, как снижается активность гормонов и ферментов, до минимума падает влажность семян (5-10 %). Покровы семени претерпевают существенные изменения: их ткани частично отмирают, уплотняются и нередко одревесневают. Такие зрелые семена способны переносить неблагоприятные условия среды и могут длительно сохранять (иногда до нескольких десятков лет) способность прорастать и давать жизнь новому растению. Состояние, в котором находятся такие зрелые семена, получило название физиологического покоя семян. В этом состоянии происходят метаболические процессы, дыхание, иногда «дозревание» зародыша, но способность к набуханию при поступлении влаги и прорастанию часто заторможена. Степень глубины физиологического покоя и его длительность неодинаковы. Семена выводятся из состояния покоя различным образом. Часть семян, особенно однолетних растений, легко набухает и прорастает уже под влиянием увлажнения (часто при этом требуется хотя бы кратковременное охлаждение). Для прорастания других семян и нормального развития проростка обязательна холодная стратификация, то есть длительное выдерживание их при пониженной температуре, во влажной среде и в условиях хорошей аэрации. Наконец, существует еще одна группа так называемых «твердосемянных» семян (у бобовых), семенная кожура которых в силу ее структурных особенностей водонепроницаема. Такие семена прорастают только после скарификации – искусственного нарушения целостности кожуры с помощью надцарапывания, перетирания с песком, ошпаривания кипятком. В природе такие семена набухают и прорастают обычно под влиянием резкой смены температурных режимов, способствующих нарушению целостности кожуры.
Прорастанием семян называют их переход от состояния покоя к вегетативному росту зародыша и формирующегося из него проростка. Прорастание начинается при оптимальном для каждого вида сочетании влажности и температуры среды, при свободном доступе кислорода. Прорастание семян сопровождается сложными биохимическими и анатомо-физиологическими процессами. При поступлении воды в семенах резко усиливается процесс дыхания, активизируются ферменты, запасные вещества переходят в легкоусвояемую, подвижную форму, образуются полирибосомы, и начинается синтез белка и других веществ. Рост зародыша обычно начинается с прорыва покровов удлиняющимся зародышевым корнем и гипокотилем в области микропилярного следа. После появления корня почечка развивается в побег, на котором развертываются настоящие листья. Иногда семядоли выносятся гипокотилем над поверхностью земли, зеленеют и выполняют функцию фотосинтезирующих органов проростка (надземное прорастание). В иных случаях они не освобождаются от покровов семени, остаются в земле и служат источником питания развивающегося проростка (подземное прорастание). В практике сельского хозяйства прорастание семян характеризуется всхожестью, то есть процентом семян, давших нормальные проростки в оптимальных для них условиях за определенный срок. Для сельскохозяйственных полевых культур этот срок равен 6-10 суткам, для древесных – 10-60.
13.6. Распространение плодов и семян
В начале XX в. шведский ботаник Р. Сернандер дал любым частям семенных растений, с помощью которых они способны расселяться, общее название диаспор (от греческого «диаспейро» – рассеиваю). В качестве диаспор могут выступать семена, плоды, соплодия, части вегетативного тела и даже целые растения. Плоды и семена, несомненно, относятся к важнейшему типу диаспор.
Существуют два основных типа распространения диаспор. Один – без содействия каких-либо внешних агентов, путем механизмов, выработанных в процессе эволюции самим растением, другой – с помощью различных внешних факторов: ветра, воды, животных, человека. Первый тип получил название автохории (от греческого «аутос» – сам, ««хорео» – иду, распространяюсь), второй – аллохории (от греческого «аллос» – другой). Растения соответственно называются автохорами и аллохорами. Плоды и семена автохоров рассеиваются сравнительно недалеко от материнского растения, обычно не больше нескольких метров от него. Группа автохорных растений разделяется на механохоры и барохоры. Плоды многих механохоров вскрываются по гнездам или створкам, причем семена из них высыпаются. Это происходит у фиалки трехцветной (Viola tricolor), видов тюльпана (Tulipa). Некоторые механохоры активно разбрасывают семена благодаря специальным приспособлениям в плодах, в основе которых повышенное осмотическое давление клеток механической ткани или, напротив, клеток основной ткани. Наиболее обычные растения такого рода – недотрога обыкновенная (Impatiens noli-tangere), экбалиум пружинистый, или бешеный огурец (Ecballium elaterium). На небольшие расстояния могут перемещаться плоды некоторых видов клевера вследствие изменения упругости зубцов чашечки при перемене влажности воздуха и почвы.
К барохорам относятся растения, обладающие тяжелыми плодами и семенами. К ним можно отнести желуди дуба (Quercus), плоды грецкого ореха (Juglans regia), семена конского каштана (Aesculus hippocastanum). При падении семена этих растений оказываются обычно под кроной материнского растения. К группе автохоров относятся также геокарпные растения (от греческого «ге» – земля). У геокарпных видов плоды в процессе развития внедряются в почву и там созревают. Наиболее известный из них арахис подземноплодный, или земляной орех (Arachis hypogaea).
Существуют четыре основных способа аллохории. Это анемохория (от греческого «анемос» – ветер), зоохория («зоон» – животное), гидрохория («гидор» – вода) и антропохория («антропос» – человек). Наиболее обширна группа растений-анемохоров. Единицы расселения, семена или целые плоды анемохоров переносятся движением воздуха. Для невскрывающихся плодов анемохоров характерны разнообразные летательные приспособления: летучки, крылатки. Классический пример растений, имеющих плоды-летучки, – одуванчик (Taraxacum officinale). Его плоды способны перелетать по воздуху на значительные расстояния. Крылатые плоды ясеня обыкновенного (Fraxinus excelsior) и тропического бобового – птерокарпуса индийского (Pterocarpus indica), оторвавшись от материнского растения, могут планировать на несколько десятков метров. Крылом, возникшим из прицветника и несущим целое соплодие, обладают виды липы (Тilia). Иногда анемохорами бывают не только растения со специально приспособленными для этого плодами, но и растения с различными летательными выростами у семян. При этом плоды обязательно вскрываются, высыпающиеся семена подхватываются ветром и разносятся. Общеизвестен тополевый «пух», который представляет снабженные волосками-летучками семена разных видов тополя (Populus), разлетающиеся из вскрывшихся плодов-коробочек.
Плоды и семена, не имеющие специальных приспособлений для переноса воздухом, тем не менее, могут разноситься на далекие расстояния путем перемещения ветром всего материнского растения, точнее, его надземной части. Речь идет об особой экологической группе растений, носящих название «перекати-поле». Эти растения, относящиеся к разным по систематическому положению семействам, подсыхая, отрываются и кочуют в виде своеобразных клубков, рассеивающих семена из вскрывшихся плодов. К растениям такого типа относятся многие степные виды – качим развесистый, или метельчатый (Gypsophila paniculata, гвоздичные), солянка холмовая (Salsola collina, маревые).
У гидрохоров плоды, распространяющиеся с помощью воды, снабжены плотным, малопроницаемым для воды эндокарпием, волокнистым легким мезокарпием, а также воздухоносными полостями, поддерживающими плод на плаву. Хорошо известны тропические гидрохоры – кокосовая пальма (Cocos nucifera) и энтада фасолевидная (Entada phaseoloides). Их плоды могут переноситься морскими и океаническими течениями на тысячи километров. Плоды пресноводных гидрохоров нередко снабжены плавательными поясками из опробковевших клеток и плотной кожурой. Большинство растений, растущих вдоль берегов прудов, озер и каналов, относятся к этой экологической группе (частуха подорожниковая Alisma plantago-aquatica, вех ядовитый Cicuta virosa).
Разнообразны способы распространения плодов и семян с помощью животных и птиц. Есть плоды, прикрепляющиеся к шерсти и ногам животных с помощью различных прицепок, крючков, шипов, а также клейкой поверхности или железистого опущения – экзозоохория. Многие плоды и семена расселяются эндозоохорно, поедаемые животными, птицами и насекомыми, разносящими семена на значительные расстояния. Классические зоохоры – виды лопуха (Аrctium), чьи соплодия, обладающие очень цепкими листочками обертки, легко переносятся шерстью животных. К лапам животных с помощью липкой поверхности прикрепляются плоды линнеи северной (Linnaea borealis). Обычный эндозоохор наших северных хвойных лесов – толокнянка. Особую группу среди зоохоров составляют растения-мирмекохоры, плоды и семена которых имеют мясистые сладкие придатки и разносятся муравьями. Антропохория среди растений тесно связана с деятельностью человека (распространение с помощью технических и транспортных средств, домашних животных). Антропохорами являются многие сорные растения.
Плоды и семена широко используются человеком для практических целей, в растениеводстве, земледелии, как продукты питания, в технике и медицине. Это связано с тем, что в них в большом количестве накапливаются различные запасные вещества – углеводы, белки и жиры, а также витамины. Кроме того, во многих плодах концентрируются вторичные метаболиты, например алкалоиды, терпеноиды, фенольные соединения. Это делает их особенно важными для медицины и фармацевтической промышленности. Особенно ценны плоды культивируемых растений, значительно более продуктивных, чем их дикорастущие сородичи. Семена и плоды – основной посевной материал. Некоторые плоды и семена применяют для изготовления бытовых украшений (бус, ожерелий, четок), а также предметов одежды (пуговицы), посуды и игрушек. Многие плоды и семена, оказавшиеся погребенными в отложениях осадочных пород, характеризуют флору и растительность прошлых геологических эпох и используются для их реконструкции. Плоды и семена растений, найденные археологами при раскопках, служат основой для воссоздания особенностей ушедших в прошлое человеческих цивилизаций.