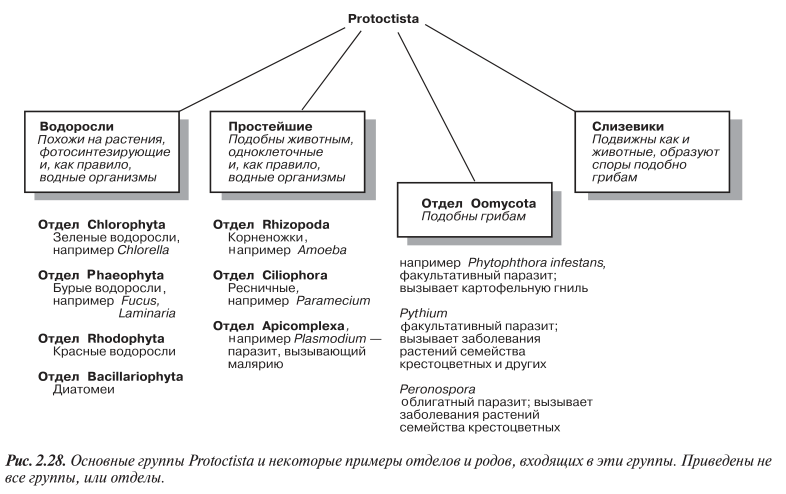
Протоктисты. Царство Protoctista
Биология — Тейлор Д., Грин Н., Стаут У. 1 том — 2013
От греч. protos — самый первый; ktistos — основать, устанавливать
2.6.1. Систематика и свойства протоктистов
Cреди царств в систематике Маргелиса и Шварца Protoctista, возможно, является самой спорной группой, поскольку естественные связи между входящими в нее организмами очень слабы. По сути в эту группу собраны все эукариотические организмы, не укладывающиеся по строгим критериям ни в одно из эукариотических царств. Многие из протоктистов — одноклеточные организмы. В царство Protoctista входят эукариоты, которые обычно считаются идентичными или подобными предкам современных растений, животных и грибов (рис. 2.28). Сюда же относятся и организмы, напоминающие ранние растения, (водоросли), ранних животных (простейшие) и ранние грибы (Oomycota). Кроме того, это царство включает и группу, известную как слизевики, которые подобно грибам образуют споры, но при этом способны «ползти» по поверхности, т. е. подобно животным они подвижны. Самые ранние эукариоты были, вероятно, одноклеточными организмами, передвигавшимися с по мощью жгутиков.
Протоктисты самая притягательная группа для тех, кто интересуется эволюцией, поскольку эти организмы служат звеном между прокариотами и более современными эукариотами, такими как растения и животные. В шестидесятые годы XX в., например, было обнаружено, что митохондрии — эти силовые станции клеток, вырабатывающие энергию в процессе аэробного дыхания, — содержат собственные ДНК и рибосомы, напоминающие ДНК и рибосомы прокариот. В результате изучения последовательности оснований в митохондриальной ДНК были получены весьма убедительные доводы в пользу того, что прежде митохондрии были аэробными бактериями (прокариотами), поселившимися некогда в предковой эукариотической клетке и «научившимися» симбиотически жить в ней. Теперь митохондрии имеются во всех эукариотических клетках и «вести» независимую жизнь они уже не способны.
Подобно митохондриям, хлоропласты — содержащие хлорофилл органеллы, — в которых происходит фотосинтез, также имеют свои собственные прокариотические ДНК и рибосомы.
По-видимому, хлоропласты произошли от фотосинтезирующих бактерий, поселившихся в свое время в подобных животным гетеротрофных клетках, превратив их в автотрофные водоросли.
Точно так же, вполне возможно, что красные водоросли произошли от сине-зеленых бактерий (цианобактерий), а зеленые водоросли — от зеленых бактерий, известных как прохлорофиты.
Теория, согласно которой митохондрии и хлоропласты являются потомками симбиотических бактерий, получила название эндосимбиотической теории. Эндосимбионт — это организм, симбиотически живущий внутри (endo-) другого организма.
2.6.2. Отдел Oomycota
Оомицеты (Oomycota) — близкие родичи грибов и схожи с ними по своему строению, однако, согласно современным представлениям, это более древняя группа. Опорным материалом клеточной стенки у оомицет служит целлюлоза, а не хитин, как у грибов. Гифы у них не разделены перегородками (нечленистые). К оомицетам относится целый ряд патогенных грибов, включая возбудителя ложной мучнистой росы — мильдью. В качестве примера рассмотрим один из таких паразитических грибов — Phytophthora infestans, который обычно считают облигатным паразитом. Для сравнения рассмотрим и другой облигатный паразит — Peronospora. И, наконец, в качестве типичного факультативного паразита рассмотрим Pythium. К облигатным паразитам относятся организмы, способные выживать и развиваться исключительно на живых клетках, в отличие от факультативных паразитов, которые обычно вызывают гибель своих хозяев, а потом живут сапротрофно на мертвых остатках.
Phytophthora infestans
Phytophthora infestans — патогенный гриб, имеющий важное хозяйственное значение, так как он паразитирует на картофеле и опустошает поля, вызывая очень опасное заболевание, известное под названием «картофельная гниль». P. Infestans не способна расти независимо от своего хозяина, напоминая в этом отношении облигатных паразитов. По своему строению и способу инфицирования фитофтора похожа на Peronospora— еще одного представителя Oomycota, являющегося возбудителем довольно распространенного, хотя и менее опасного заболевания желтофиоли, капусты и многих других членов семейства крестоцветных (Crucifera).
Мицелий фитофторы, перезимовав в клубнях картофеля, весной начинает расти на листьях растения, где обычно в августе появляются первые видимые признаки гнили.
Мицелий, состоящий из разветвленных нечленистых гиф, распространяется по межклеточным пространствам внутри листьев, образуя ветвящиеся гаустории, которые, проникая в клетки мезофилла, высасывают из них питательные вещества (рис. 2.29).
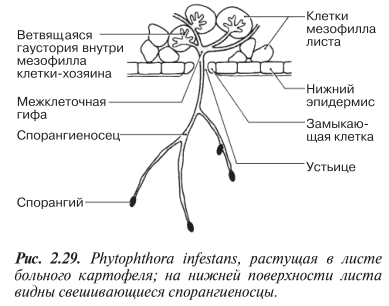
Гаустории — типичные для облигатных паразитов образования специализированы для проникновения и всасывания.
Каждая из гаусторий представляет собой видоизмененный вырост гифы, обладающий большой поверхностью. Такие выросты (гаустории) проникают в клетки, не разрушая плазматические мембраны и не убивая клетки. В теплой и влажной среде мицелий образует спорангиеносцы, выступающие из нижней стороны листа через устьица или раны. Спорангиеносцы ветвятся, давая начало спорангиям (рис. 2.29). В теплой среде спорангии могут вести себя как споры, т. е. переноситься ветром или с брызгами от капель
дождя на другие растения, распространяя таким образом инфекцию. Затем из спорангия вырастает гифа, которая проникает через устьица, чечевички или повреждения внутрь ткани растения. В холодных условиях содержимое спорангия делится с образованием подвижных зооспор (этот признак характерен для примитивных организмов), которые высвобождаются из спорангия и плавают в тонком слое жидкости, адсорбированной на поверхности листа. Зооспоры могут инцистироваться и в таком состоянии дожидаться, пока условия не станут более благоприятными для роста гиф; тогда-то и начинается заражение новых растений.
У больных растений на отдельных листьях видны небольшие мертвые, «гнилые» зоны коричневого цвета. Если присмотреться повнимательнее, то на нижней поверхности зараженных листьев вокруг мертвой зоны можно разглядеть бахрому из белых спорангиеносцев. При теплой сырой погоде зоны некроза быстро распространяются по всей поверхности листа и переходят на стебель. Некоторые спорангии падают на землю и заражают клубни картофеля, при этом инфекция распространяется очень быстро и вызывает своего рода сухую гниль, при которой ткань клубня приобретает ржаво-коричневую окраску, неравномерно распространяющуюся от периферии к его центру.
Сначала корневая шейка, а затем и все остальные части растения превращаются в гнилую жижу, так как зоны некроза вторично инфицируются сапротрофными бактериями-редуцентами. Таким образом, фитофтора совсем убивает растение, что отличает ее от ближайшего родича —
Peronospora, являющегося облигатным паразитом. В этом отношении Phytophthoraне похожа на типичного облигатного паразита, и иногда ее относят к факультативным паразитам, хотя здесь, пожалуй, не стоит особо останавливаться на таких нюансах.
Фитофтора, как правило, зимует в состоянии спящего мицелия внутри слегка инфицированных клубней картофеля. Считается, что в отличие от Peronospora этот гриб редко размножается половым путем, если, конечно, не говорить о тех местах (Мексике, Центральной и Южной Америке), откуда произошел картофель. Половое размножение гриба можно индуцировать в лабораторных условиях. Как и Peronospora, фитофтора образует устойчивые покоящиеся споры. Толстостенная спора образуется в результате слияния антеридия (мужской репродуктивный орган) и оогония (женский репродуктивный орган). Такая спора может перезимовать в почве и на следующий год вызвать новую инфекцию.
В прошлом эпидемии, вызываемые фитофторой, приводили к очень серьезным последствиям. Полагают, что болезнь случайно завезли в Европу из Америки в конце тридцатых годов XIX в. В результате по Европе прокатилась целая волна эпидемий, которые в 1845 г. и в последующие годы полностью уничтожили посевы картофеля в Ирландии. Наступил голод, который привел к гибели многих людей, оказавшихся жертвами не только самой болезни картофеля, но и сложных политических и экономических последствий. В результате многие ирландские семьи были вынуждены эмигрировать в Северную Америку.
Эта болезнь представляет интерес и в том отношении, что в 1845 г. Беркли (Berkeley) впервые показал ее микробную природу. Беркли продемонстрировал, что гриб, связанный с картофельной гнилью, сам вызывает болезнь, а не является побочным продуктом разложения.
Выяснение жизненного цикла возбудителя картофельной гнили привело к разработке методов борьбы с этой болезнью. Ниже перечислены эти методы.
- 1. Необходимо тщательно следить за тем, чтобы не был высажен ни один зараженный клубень.
- 2. Поскольку гриб может сохраняться в почве почти целый год, не следует сажать картофель там, где было обнаружено это заболевание в предыдущем году. В этом случае целесообразны правильные севообороты.
- 3. Все больные части растений следует уничтожить еще до выкапывания клубней, например сжечь их или опрыскать каким-либо едким раствором, таким как серная кислота. Это необходимо потому, что гнилая ботва (т. е. стебли) и другие надземные части могут заразить и клубни.
- 4. Поскольку этот гриб может зимовать в невыкопанных клубнях, необходимо тщательно следить, чтобы на зараженных полях не оставалось ни одного клубня.
- 5. Гриб можно уничтожать медьсодержащими фунгицидами, например бордосской жидкостью. Опрыскивание следует проводить в строго определенное время, чтобы успеть предупредить заболевание, так как пораженное растение уже ничто не спасет. Растения обычно опрыскивают через каждые две недели, начиная с того момента, когда они вырастут до нескольких сантиметров в высоту, и до тех пор, пока они полностью не созреют. Клубни, отобранные для последующего посева, можно простерилизовать снаружи, погрузив их в разбавленный раствор хлористой ртути(II).
- 6. Постоянное наблюдение за метеорологическими условиями и раннее оповещение фермеров могут помочь определить, когда следует опрыскивать посевы.
- 7. Одно время проводили селекцию картофеля на устойчивость к гнили. Как известно, дикий картофель Solanum demissum обладает высокой устойчивостью к фитофторе, поэтому его использовали в опытах по селекции. Самое большое препятствие для получения нужного иммунитета заключается в том, что существует много штаммов этого гриба, и до сих пор не удалось вывести ни одного сорта картофеля, который был бы устойчив ко всем штаммам. По мере введения в культуру новых сортов картофеля могут появляться и новые штаммы этого гриба. Указанная проблема давно знакома фитопатологам; она лишний раз напоминает нам о необходимости сохранения генофонда диких предков наших современных сельскохозяйственных культур как источника генов устойчивости к различного рода заболеваниям.
Pythium
В отличие от фитофторы Pythium— относительно неспециализированный паразит, поражающий большое число различных растений и вызывающий мягкую гниль. Pythium вызывает «вымокание» проростков. Ему необходима влажная среда, поскольку при бесполом размножении он образует плавающие споры. Гриб этот может расти как на живом растении, так и на мертвых остатках, следовательно, это факультативный паразит.
Он может жить и сапротрофно в сырой почве. Pythium выделяет во внешнюю среду ферменты, что дает ему возможность быстро атаковать и убивать хозяина. Первыми из ферментов образуются пектиназы, которые, диффундируя впереди растущего гриба, растворяют пектин срединной пластинки, удерживающей клетки вместе. В результате ткани растения превращаются в кашу (мягкая гниль) и растение погибает. Позднее образуются другие ферменты, которые переваривают содержимое растительных клеток. Однако гаустории у Pythium в отличие от фитофторы не образуются. Продукты переваривания поглощаются гифами, растущими между клетками.
Вымокание обусловлено разрушением первых проростков по мере их появления над поверхностью почвы. Первые водянистые пятна на стебле появляются на уровне почвы. Когда они темнеют, стебель погибает. Вымокание проростков может создавать серьезную проблему в садоводстве, лесоводстве и в сельском хозяйстве.
Наиболее чувствительны к этому заболеванию представители семейства крестоцветных, особенно когда проростки растут очень густо.
2.6.3. Водоросли
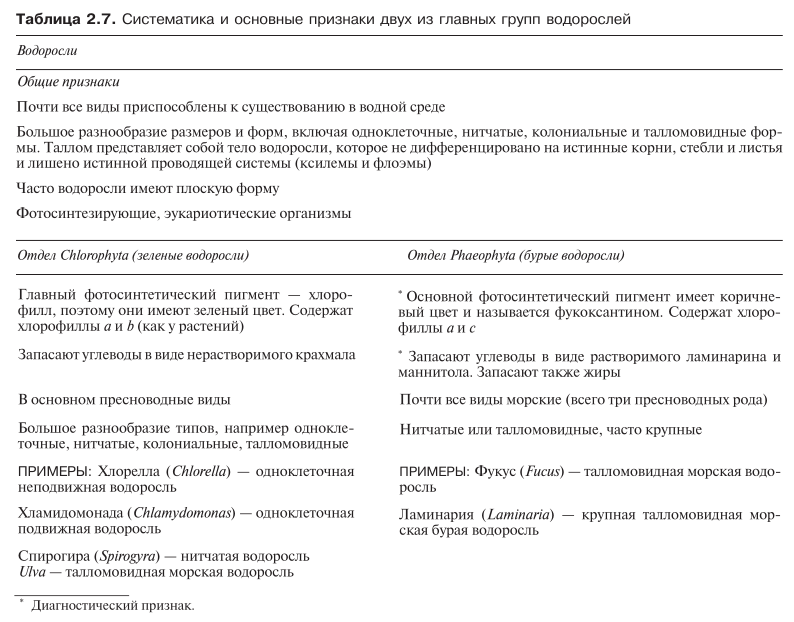
Водоросли образуют огромную группу протоктистов, имеющих большое биологическое значение и очень важных для человека. У них нет ни одного диагностического признака. Вероятнее всего, что это фотосинтезирующие эукариоты, которые эволюционировали в водной среде и в ней остались. Некоторые водоросли вышли из воды и успешно приспособились к жизни на суше, но в отличие от растений доля наземных водорослей ничтожна по сравнению с океаническими и пресноводными формами. Тело водорослей не разделяется на стебель, корни и листья. Такое относительно недифференцированное тело называют талломом.
Водоросли образуют ряд четко выраженных естественных групп, различающихся главным образом своими фотосинтетическими пигментами. В современной классификации эти группы получили статус отделов. На рис. 2.28 приводятся только четыре отдела водорослей. Характерные особенности водорослей в целом, а также двух из основных отделов описаны в табл. 2.7. Два представителя водорослей, а именно Chlorella (отдел Chlorophyta) и Fucus (отдел Phaeophyta) рассматриваются ниже более подробно.
2.6.4. Отдел Chlorophyta (зеленые водоросли)
Хлорелла
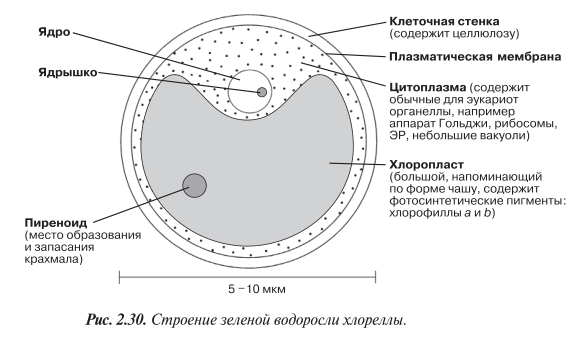
Хлорелла — одноклеточная, неподвижная зеленая водоросль. Ее строение показано на рис. 2.30. Ее можно встретить в пресноводных прудах и канавах. Хлореллу легко культивировать и она широко используется в экспериментах по изучению фотосинтеза (разд. 7.6), а также в качестве альтернативного источника питания (белок одноклеточных).
2.6.5. Отдел Phaeophyta (бурые водоросли)
Фукус
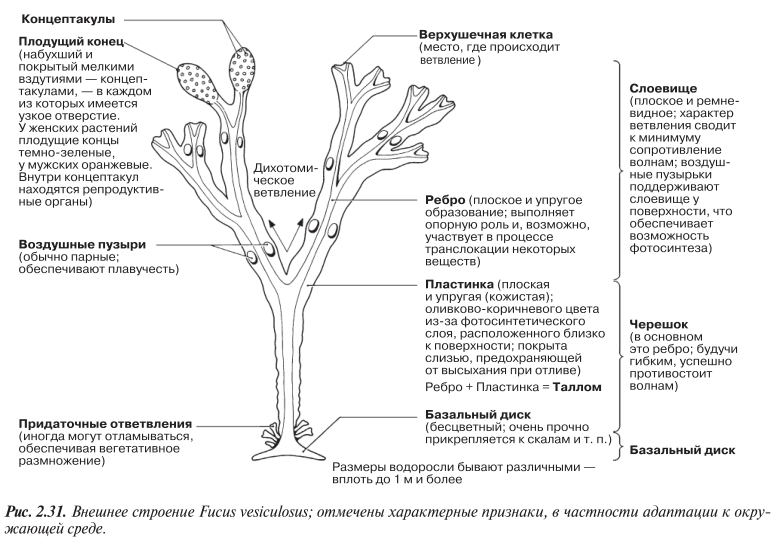
Фукус — относительно крупная бурая водоросль с довольно сложным строением. Ее тело представляет собой таллом, дифференцированный на черешок, базальный диск и слоевище (следует иметь в виду, что это не настоящие стебель, корни и листья). Эта морская водоросль часто встречается у скалистых берегов Британского побережья. Она хорошо адаптирована к суровым условиям побережья, где из-за приливов и отливов попеременно то обнажается, то вновь покрывается водой.
Известны три наиболее распространенных вида фукуса, которые часто встречаются в трех разных зонах, или глубинах, побережья — явление, называемое зональным распределением. Распределение водорослей по зонам связано главным образом с их способностью выдерживать пребывание на воздухе. Ниже перечислены основные признаки, по которым их можно узнать, а также места их распространения на побережье.
F. spiralis (плоская водоросль) — их выбрасывает на берег у самой высокой отметки прилива. В погруженном состоянии таллом слегка закручен в спираль.
F. serratus (обыкновенная зубчатая или пильчатая водоросль) — в средней приливной зоне. Края таллома зазубрены.
F. vesiculosus (пузырчатая водоросль) — у самой низкой отметки отлива. Имеются воздушные пузыри, обусловливающие плавучесть водоросли. Внешние признаки F. vesiculosus показаны на рис. 2.31.
АДАПТАЦИИ К ОКРУЖАЮЩЕЙ СРЕДЕ.
Прежде чем мы рассмотрим адаптации фукуса к среде обитания, следует сказать несколько слов о природе самой среды, которая достаточно негостеприимна. Будучи растениями приливно-отливной зоны, водоросли разных видов в разной степени подвергаются воздействию воздушной среды во время отлива. Поэтому они должны быть защищены от высыхания. Кроме того, и температура может резко меняться, когда холодные морские волны вливаются в прогретые лужицы, оставшиеся после отлива. Растения должны быть адаптированы и еще к одному фактору, а именно к резким
изменениям солености воды, будь то ее увеличение при испарении из небольших водоемов, образовавшихся после отлива, или ее уменьшение во время дождя. Для того чтобы противостоять таким факторам, как приливы, отливы, прибой и удары волн, нужна определенная механическая прочность. Большие волны способны перекатывать камни, которые могут придавливать водоросли, нанося им большие повреждения.
МОРФОЛОГИЧЕСКИЕ АДАПТАЦИИ (ОБЩЕЕ СТРОЕНИЕ).
Таллом водоросли прочно прикрепляется к грунту с помощью базального диска (рис. 2.31). Связь с субстратом, обычно это камни, оказывается настолько прочной, что водоросль бывает чрезвычайно трудно оторвать от него. На деле первым, как правило, не выдерживает камень, а не базальный диск.
Таллом дихотомически ветвится (т. е. образует по две ветви в каждой точке ветвления). Это сводит к минимуму сопротивление потоку воды, устремляющейся между ветвями. К тому же таллом прочный и упругий, но не жесткий, а ребра слоевища прочные и гибкие.
У F. vesiculosus имеются воздушные пузыри, обеспечивающие его плавучесть; это удерживает слоевище вблизи поверхности воды, т. е. в условиях, способствующих максимальному улавливанию света для фотосинтеза.
Хлоропласты расположены в поверхностных слоях водоросли, обусловливая максимальное воздействие необходимого для фотосинтеза света.
ФИЗИОЛОГИЧЕСКИЕ АДАПТАЦИИ
Среди фотосинтетических пигментов преобладает бурый пигмент фукоксантин. В этом проявляется адаптация к фотосинтезу под водой, поскольку фукоксантин сильно поглощает свет в синей области видимого спектра, проникающий в толщу воды гораздо дальше, чем свет с большей длиной волны, например красный.
Таллом секретирует в больших количествах слизь, заполняющую все внутренние полости водоросли и выделяющуюся на поверхность. Слизь помогает удерживать воду, препятствуя таким образом обезвоживанию во время отлива. Осмотический потенциал клеток водоросли выше (менее отрицательный), чем осмотический потенциал морской воды, и поэтому потерь воды за счет осмоса здесь не происходит.
ПРИСПОСОБЛЕНИЯ К ПОЛОВОМУ РАЗМНОЖЕНИЮ
Высвобождение гамет у фукуса синхронизировано с приливами. Во время отлива таллом обсыхает и выдавливает наружу репродуктивные органы, защищенные от высыхания слизью. Во время прилива стенки репродуктивных органов растворяются, высвобождая гаметы.
Мужские гаметы подвижны и обладают положительным хемотаксисом, обусловливающим их перемещение в сторону химического секрета, выделяемого женскими гаметами. Развитие зиготы происходит сразу после оплодотворения, что сводит к минимуму риск быть смытым в океан.
2.6.6. Простейшие
Как и водоросли простейшие образуют большую группу протоктистов. Это одноклеточные, подобные животным организмы с гетеротрофным способом питания. Существуют свыше 5 известных видов, которые можно обнаружить всюду, где есть вода. Большинство из них — свободно живущие организмы, характеризующиеся различными способами передвижения. Некоторые, однако, являются паразитами. Это, в частности, Plasmodium, вызывающий болезнь, которая, по имеющимся оценкам, убила больше людей, чем любая другая. Эта болезнь — малярия — все еще остается одной из страшнейших в мире убийц. В этом разделе в качестве примера для изучения простейших мы выбрали свободно живущий организм инфузорию туфельку (Paramecium caudatum), поскольку она типична для этого уровня организации. Малярийный плазмодий рассматривается в гл. 15.
2.6.7. Отдел Ciliophora (ресничные)
Ресничные — один из типов простейших (рис. 2.28). Им присущи следующие признаки:
- 1) это одноклеточные гетеротрофные формы;
- 2) они снабжены ресничками — тонкими волосковидными отростками, биения которых вызывают ток воды, используемый клеткой либо для своего передвижения, либо для захвата пищевых частиц;
- 3) для них характерна определенная форма клетки, обусловленная наличием тонкого гибкого наружного слоя цитоплазмы, называемого пелликулой. Пелликула в свою очередь окружена плазматической мембраной;
- 4) клетки ресничных имеют довольно сложное строение, например они содержат макронуклеус и микронуклеус.
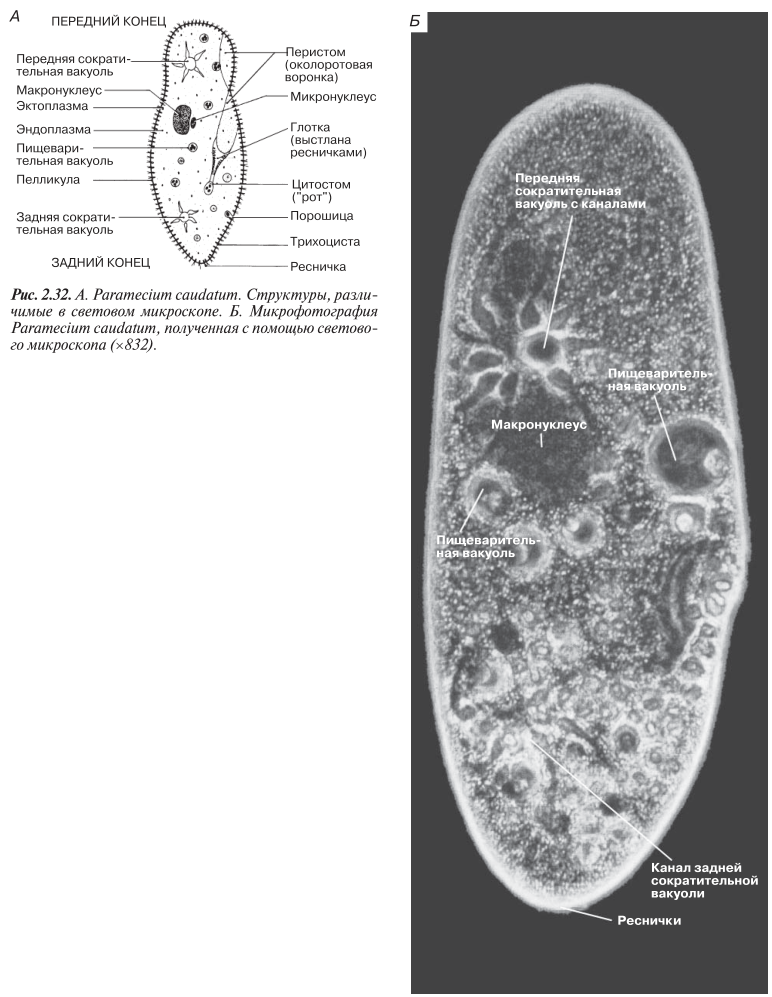
Наиболее типичный широко распространенный представитель ресничных — инфузория туфелька (Paramecium). Она обитает в стоячей воде, а также в пресноводных водоемах с очень слабым течением, содержащих разлагающийся органический материал. Рис. 2.32 дает представление о довольно сложном строении этих организмов, типичном для инфузории. Сложность строения клетки у парамеции объясняется тем обстоятельством, что ей приходится выполнять все функции, присущие целому организму, а именно питание, осморегуляцию и передвижение. Тело парамеции имеет характерную форму: передний конец у нее тупой, а задний несколько заострен. Реснички расположены парами по всей поверхности клетки. Располагаясь продольными диагональными рядами, они, совершая биения, заставляют инфузорию вращаться и продвигаться вперед. Между ресничками находятся отверстия, ведущие в особые камеры, называемые трихоцистами.
Из этих камер под влиянием определенных раздражителей могут выстреливать тонкие остроконечные нити, используемые, вероятно, для удержания добычи. Под пелликулой располагается эктоплазма — прозрачный слой плотной цитоплазмы консистенции геля. В эктоплазме находятся базальные тельца (идентичные центриолям), от которых отходят реснички, а между базальными тельцами имеется сеть тонких фибрилл, участвующих, по-видимому, в координировании биения ресничек. Основная масса цитоплазмы представлена эндоплазмой, имеющей более жидкую консистенцию, чем эктоплазма. Именно в эндоплазме расположено большинство органелл. На вентральной (нижней) поверхности туфельки ближе к ее переднему концу находится околоротовая воронка, на дне которой находится рот, или цитостом. Рот ведет в короткий канал — цитофаринкс, или глотку. Как околоротовая воронка, так и глотка могут быть выстланы ресничками, движения которых направляют к цитостому поток воды, несущей с собой различные пищевые частицы, такие, например, как бактерии.
Вокруг попавших в цитоплазму путем эндоцитоза пищевых частиц образуется пищевая вакуоль. Эти вакуоли перемещаются по эндоплазме к так называемой порошице, через которую непереваренные остатки путем экзоцитоза выводятся наружу. В цитоплазме имеются также две сократительные вакуоли, местоположение которых в клетке строго фиксировано (рис. 2.32). Эти вакуоли отвечают за осморегуляцию, т. е. поддерживают в клетке определенный водный потенциал. Жизнь в пресной воде осложняется тем, что в клетку постоянно поступает вода в результате осмоса; эта вода должна непрерывно выводиться из клетки, чтобы предотвратить ее разрыв. Происходит это с помощью процесса активного транспорта, требующего затраты энергии. Вокруг каждой сократительной вакуоли расположен ряд расходящихся лучами каналов, собирающих воду, перед тем как высвободить ее в центральную вакуоль. В клетке парамеции находятся два ядра. Большее из них — макронуклеус— полиплоидное; оно имеет более двух наборов хромосом и контролирует метаболические процессы, не связанные с размножением. Микронуклеус — диплоидное ядро. Оно контролирует размножение и образование макронуклеусов при делении ядра. Парамеция может размножаться и бесполым путем (поперечным делением надвое) и половым (путем конъюгации).
2.6.8. Отдел Apicomplexa
У простейших этой группы также имеется пелликула, придающая клетке определенную форму. Однако у большинства из них нет специальных структур для передвижения. Отличительная особенность Apicomplexa — это образование спор и при половом и при бесполом размножении. Примером этих простейших служит паразит Plasmodium — возбудитель малярии у человека