Общая характеристика растений. Споровые растения
Ботаника. Учебное пособие по специальности «Фармация»
Царство растения – Plantae, Vegetabilia
Общая характеристика царства
Представители царства – высокоспециализированные автотрофные организмы, питающиеся за счет процесса аэробного фотосинтеза. Их тело обычно расчленено на стебель, корень, лист, и они хорошо приспособлены к жизни в наземно-воздушной среде. Клетки растений имеют плотную клеточную стенку, в основе которой – целлюлоза. Основной запасной продукт – крахмал. Размножение вегетативное, бесполое (спорами) и половое (оогамия); мужские гаметы либо имеют ундулиподии (сперматозоиды), либо не имеют их (спермии). Характерно чередование полового (гаметофит) и бесполого поколения (спорофит) с преобладанием диплоидного бесполого поколения. Зигота у растений дает начало зародышу, который затем развивается в спорофит.
Царство растений включает не менее 300 тыс. видов (ныне существующих и вымерших), относящихся к 9 отделам – риниофиты (Rhyniophyta) и зостерофиллофиты (Zosterophyllophyta) (ныне вымершие), моховидные (Btyophyta), плауновидные (Lycopodiophyta), псилотовидные (Psilotophyta), хвощевидные (Equisetophyta), папоротниковидные (Polypodiophyta), голосеменные (Pinophyta) и покрытосеменные (Magnoliophyta). Представители ныне существующих отделов, за исключением моховидных, характеризуются преобладанием в цикле развития бесполого поколения (спорофита), имеющего сосуды и (или) трахеиды. Благодаря последнему обстоятельству эти растения называют сосудистыми.
Растения делят на две группы: споровые и семенные. У споровых растений спорогенез и гаметогенез разобщены во времени и пространстве: спорофиты и гаметофиты являются отдельными физиологически самостоятельными организмами. Единицей размножения являются споры. У семенных растений гаметофиты сильно редуцированы и не являются физиологически самостоятельными организмами. Единицей размножения является семя.
Споровые растения являются первопоселенцами суши, которые дали начало в процессе эволюции семенным растениям.
Споровые растения
Включают следующие ныне существующие отделы: моховидные (Bryophyta), плауновидные (Lycopodiophyta), псилотовидные (Psilotophyta), хвощевидные (Equisetophyta), папоротниковидные (Polypodiophyta).
Споровые растения появились в конце силурийского периода, более 400 млн. лет тому назад. Первые представители споровых были небольших размеров и имели простое строение, но уже у примитивных растений наблюдалась дифференциация на элементарные органы. Усовершенствованию органов соответствовало усложнение внутреннего строения и онтогенеза. В жизненном цикле происходит чередование полового и бесполого способов размножения и связанное с этим чередование поколений. Бесполое поколение представлено диплоидным спорофитом, половое – гаплоидным гаметофитом.
На спорофите образуются спорангии, внутри которых в результате мейотического деления образуются гаплоидные споры. Это мелкие, одноклеточные образования, лишенные жгутиков. Растения, у которых все споры одинаковые, называются равноспоровыми. У более высокоорганизованных групп споры двух типов: микроспоры (формируются в микроспорангиях), мегаспоры (формируются в мегаспорангиях). Это разноспоровые растения. При прорастании споры образуется гаметофит.
Полный жизненный цикл (от зиготы до зиготы) состоит из гаметофита (период от споры до зиготы) и спорофита (период от зиготы до образования спор). У плаунов, хвощей и папоротников эти фазы представляют собой как бы отдельные физиологически самостоятельные организмы. У мхов гаметофит является самостоятельной фазой жизненного цикла, а спорофит сведен до его своеобразного органа – спорогона (спорофит живет на гаметофите).
На гаметофите развиваются органы полового размножения: архегонии и антеридии. В архегониях, похожих на колбу, образуются яйцеклетки, а в мешковидных антеридиях — сперматозоиды. У равноспоровых растений гаметофиты обоеполые, у разноспоровых – однополые. Оплодотворение происходит лишь при наличии воды. При слиянии гамет образуется новая клетка – зигота с двойным набором хромосом (2n).
Отдел Моховидные – Bryophyta
Насчитывает до 27000 видов. Моховидные имеют тело либо в виде слоевища, либо расчленены на стебель и листья. Настоящих корней у них нет, их заменяют ризоиды. Проводящие ткани появляются лишь у высокоразвитых мхов. Частично обособлены ассимиляционная и механическая ткани.
В жизненном цикле доминирует гаметофит. Спорофит самостоятельно не существует, развивается и всегда находится на гаметофите, получая от него воду и питательные вещества. Спорофит представляет собой коробочку, где развивается спорангий, на ножке, связывающей ее с гаметофитом.
Размножаются мхи спорами, могут размножаться и вегетативным путем – отдельными участками тела или особыми выводковыми почками.
Отдел делится на три класса: Антоцеротовые (100 видов, шесть родов талломных растений), Печеночные и Листостебельные мхи.
Класс Печеночные мхи (Hepaticopsida)
Класс насчитывает около 8500 видов. Это, в основном, слоевищные мхи, хотя есть виды, имеющие стебель и листья. Широко распространена маршанция обыкновенная (Marchantia polymorpha) (рис 11. 1).
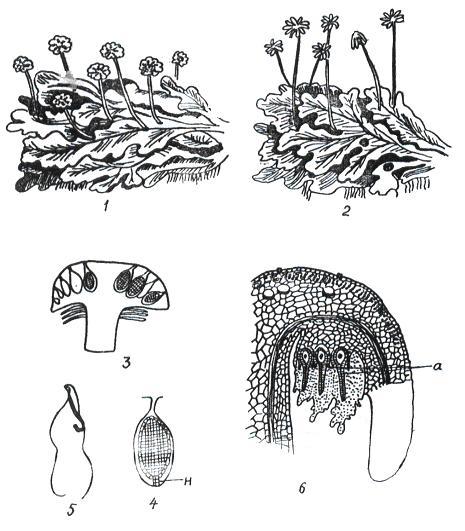
Рис. 11. 1. Цикл воспроизведения маршанции: 1 – таллом с мужскими подставками; 2 – таллом с женскими подставками; 3 – вертикальный разрез через мужскую подставку (в некоторых антеридиальных полостях находятся антеридии); 4 – антеридий в антеридиальной полости (н – ножка антеридия); 5 – двужгутиковый сперматозоид; 6 – вертикальный разрез через женскую подставку (а – архегоний).
Гаметофит имеет темно-зеленое слоевище (таллом), дихотомически разветвленное на широкие лопастные пластинки с дорсовентральной (спинно-брюшной) симметрией. Сверху и снизу слоевище покрыто эпидермой, внутри имеются ассимиляционная ткань и клетки, выполняющие проводящую и запасающую функции. К субстрату слоевище прикрепляется ризоидами. На верхней стороне слоевища в особых «корзиночках» образуются выводковые почки, служащие для вегетативного размножения.
Талломы раздельнополые, органы полового размножения развиваются на особых вертикальных ветвях-подставках.
Мужские гаметофиты имеют восьмилопастные подставки, на верхней стороне которых находятся антеридии. На женских гаметофитах подставки со звездчатыми дисками, на нижней стороне лучей звездочки расположены (шейкой вниз) архегонии. При наличии воды сперматозоиды перемещаются, попадают в архегоний и сливаются с яйцеклеткой.
После оплодотворения из зиготы развивается спорогон. Он имеет вид шаровидной коробочки на короткой ножке. Внутри коробочки, в результате мейоза, из спорогенной ткани, образуются споры. В благоприятных условиях споры прорастают, из них развивается протонема в виде небольшой нити, из верхушечной клетки которой развивается таллом маршанции.
Класс Листостебельные мхи (Bryopsida, или Musci).
Листостебельные мхи распространены по всему земному шару, особенно в условиях холодного климата на сырых местах, в сосновых и еловых лесах, в тундре. На торфяных и моховых болотах часто образуют плотный ковер. Тело расчленено на стебель и листья, но настоящих корней нет, есть многоклеточные ризоиды. Класс состоит из трех подклассов: Бриевые, или Зеленые мхи; Сфагновые, или Белые мхи; Андреевые, или Черные мхи.
Андреевые мхи (три рода, 90 видов) распространены в холодных областях, внешне сходны с зелеными, по строению листьев и коробочки – со сфагновыми.
Подкласс Бриевые, или Зеленые мхи (Bryidae). Насчитывает около 700 родов, объединяющих 14000 видов, широко распространенных повсюду, особенно в тундровой и лесной зонах Северного полушария.
Широко распространен кукушкин лен (Polytrichium commune), образующий густые дерновины на сырых почвах в лесах, на болотах и лугах. Стебли до40 см высоты, неветвистые, с густыми жесткими и острыми листьями. От нижней части стебля отходят ризоиды.
Цикл развития кукушкина льна (рис. 11. 2).
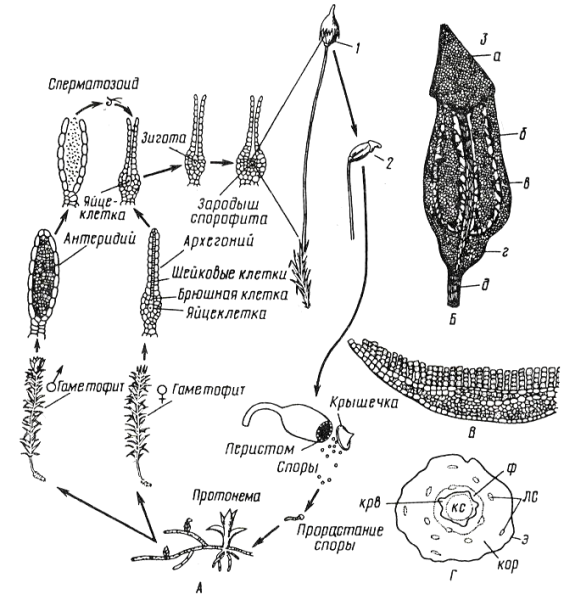
Рис. 11. 2. Кукушкин лен: А – цикл развития мха; Б – коробочка: 1 – с колпачком, 2 – без колпачка, 3 – в разрезе (а –крышечка, б – урночка, в – спорангий, г – апофиза, д – ножка); В – поперечный разрез листа с ассимиляторами; Г – поперечный разрез стебля (ф – флоэма, крв – крахмалоносное влагалище, кор – кора, э – эпидерма, лс – листовые следы).
Гаметофиты кукушкина льна раздельнополые. Рано весной на верхушке мужских особей развиваются антеридии, на верхушках женских – архегонии.
Весной во время дождя или после росы сперматозоиды выходят из антеридия и проникают в архегоний, где сливаются с яйцеклеткой. Из зиготы здесь же, на верхушке женского гаметофита, вырастает спорофит (спорогон), имеющий вид коробочки на длинной ножке. Коробочка покрыта волосистым колпачком (калиптра) (остатком архегония). В коробочке – спорангий, где после мейоза образуются споры. Спора – небольшая клетка с двумя оболочками. На верхушке коробочки, по ее краю, расположены зубцы (перистом), которые, в зависимости от влажности воздуха, загибаются внутрь коробочки или отгибаются наружу, чем способствуют рассеиванию спор. Споры разносятся ветром и в благоприятных условиях прорастают, образуя протонему. Через некоторое время на протонеме образуются почки, из которых формируются листостебельные побеги. Эти побеги вместе с протонемой – гаплоидное поколение – гаметофит. Коробочка на ножке – диплоидное поколение – спорофит.
Подкласс Сфагновые, или Белые мхи (Sphagnidae)
К сфагновым мхам относятся свыше 300 видов единственного рода сфагнум (Sphagnum) (рис. 11. 3).
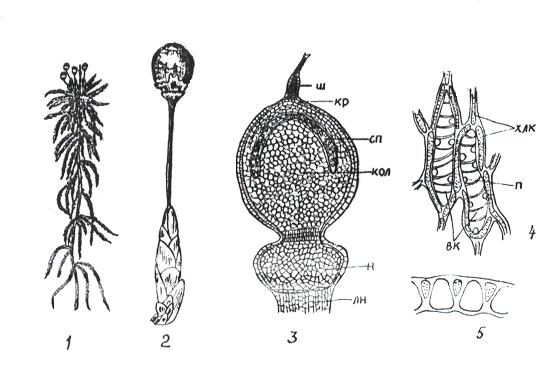
Рис 11. 3. Сфагнум: 1 – внешний вид; 2 – верхушка ветви со спорогоном; 3 – спорогон (ш – остаток шейки архегония, кр – крышечка, сп – спорангий, кол – колонка, н – ножка спорогона, лн – ложная ножка); 4 – часть веточного листа (хлк – хлорофиллоносные клетки, вк – водоносные клетки, п – поры); 5 – поперечный разрез листа.
Ветвистые стебли сфагнума усеяны мелкими листьями. На верхушке главной оси боковые веточки образуют розетку почковидной формы. Особенностью сфагновых мхов является непрерывное нарастание стебля верхушкой и отмирание нижней части. Ризоиды отсутствуют, и поглощение воды с минеральными веществами происходит стеблями. Листья этих мхов состоят из двух типов клеток: 1) живых ассимилирующих, длинных и узких, хлорофиллоносных; 2) гиалиновых – мертвых, лишенных протопласта. Гиалиновые клетки легко заполняются водой и долго ее сохраняют. Благодаря такому строению, сфагновые мхи могут накапливать воды в 37 раз больше своей сухой массы. Разрастаясь плотными дернинами, сфагновые мхи способствуют заболачиванию почв. На болотах наслаивание отмерших частей мха приводит к образованию торфяников. Путем сухой перегонки из торфа получают воск, парафин, фенолы, аммиак; путем гидролиза – спирт. Торфяные плиты являются хорошим теплоизоляционным материалом. Сфагновые мхи обладают бактерицидными свойствами.
Отдел Плауновидные – Lycopodiophyta
Появление плауновидных связывают с силурийским периодом палеозойской эры. В настоящее время отдел представлен травянистыми растениями со стелющимися, дихотомически ветвящимися стеблями и корнями, а также спирально расположенными чешуйчатыми листьями. Листья возникли как выросты на стебле, и их называют микрофиллы. У плаунов есть флоэма, ксилема, перицикл.
Выделяют два современных класса: равноспоровые Плауновые и разноспоровые Полушниковые.
Класс Плауновые (Lycopodiopsida)
Из всего класса до настоящего времени сохранилось четыре рода.
Род плаун (Lycopodium). К этому роду относят многочисленные (около 200 видов) многолетние вечнозеленые травы, распространенные от арктических областей до тропиков. Так плаун булавовидный (L. clavatum) встречается в травяном покрове хвойных лесов на достаточно влажных, но бедных гумусом почвах. В сырых хвойных лесах широко распространен плаун годичный (L. annotinum) (рис. 11. 4).

Рис. 11. 4. Плаун булавовидный.
Род баранец (Huperzia). Представитель рода — баранец обыкновенный (H. selago) распространен в тундровой, лесотундровой и северной лесной зонах и произрастает в южно-таежных еловых лесах и ольшаниках, а также в мшистых лесах и на альпийских лугах.
Род дифазиаструм (Diphasiastrum). Представитель рода дифазиаструм сплюснутый (D. complanatum) растет на сухих песчаных почвах в сосновых лесах.
Цикл развития на примере плауна булавовидного (рис. 11. 5).
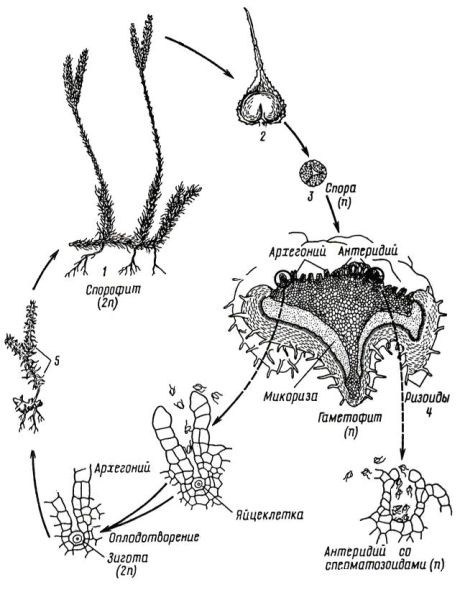
Рис. 11. 5. Цикл развития плауна булавовидного: 1 – спорофит; 2 – спорофилл со спорангием; 3 – спора; 4 – гаметофит с антеридиями и архегониями; 5 – развивающийся на гаметофите из зародыша молодой спорофит.
Стелющиеся побеги плауна булавовидного достигают до 25 см в высоту и более 3 м в длину. Стебли покрыты спирально расположенными ланцетно-линейными мелкими листьями. В конце лета на боковых побегах образуется обычно два спороносных колоска. Каждый колосок состоит из оси и мелких тонких спорофиллов – видоизмененных листьев, в основании которых расположены спорангии почковидной формы.
В спорангиях после редукционного деления клеток спорогенной ткани образуются одинаковой величины, одетые толстой желтой оболочкой гаплоидные споры. Они прорастают после периода покоя через 3-8 лет в обоеполые заростки, представляющие собой половое поколение и живущие сапротрофно в почве, в виде клубенька. От нижней поверхности отходят ризоиды. Через них в заросток врастают гифы гриба, образуя микоризу. В симбиозе с грибом, который обеспечивает питание, живет заросток, лишенный хлорофилла и неспособный к фотосинтезу. Заростки многолетние, развиваются очень медленно, лишь через 6-15 лет на них образуются архегонии и антеридии. Оплодотворение совершается при наличии воды. После оплодотворения яйцеклетки двужгутиковым сперматозоидом образуется зигота, которая без периода покоя прорастает в зародыш, развивающийся во взрослое растение.
В официнальной медицине споры плаунов применялись в качестве детской присыпки и обсыпки для пилюль. Побеги баранца обыкновенного используют для лечения больных, страдающих хроническим алкоголизмом.
Класс Полушниковые (Isoetopsida)
Селагинелла (Selaginella) среди современных родов насчитывает самое большое (около 700) число видов.
Это нежное многолетнее травянистое растение, требующее высокой влажности. Селагинеллы в отличие от плаунов характеризуются разноспоровостью. В спороносных колосках образуется два вида спор – четыре мегаспоры в мегаспорангиях и многочисленные микроспоры в микроспорангиях. Из микроспоры образуется мужской гаметофит, состоящий из одной ризоидальной клетки и антеридия со сперматозоидами. Мегаспора развивается в женский гаметофит, не покидающий ее оболочки и состоящий из мелкоклеточной ткани, в которую погружены архегонии. После оплодотворения из яйцеклетки развивается зародыш, и затем новый спорофит.
Отдел Хвощевидные – Equisetophyta
Появились хвощи в верхнем девоне, достигли наибольшего разнообразия в карбоне, когда древесный ярус заболоченных тропических лесов в значительной степени состоял из древовидных хвощевидных, вымерших к началу мезозоя. Современные хвощевидные появились на Земле с мелового периода.
До настоящего времени сохранился лишь один род – хвощ (Equisetum), представленный 30-35 видами, распространенными на всех континентах.
У всех видов хвоща стебли имеют членистое строение с выраженным чередованием узлов и междоузлий. Листья редуцированы до чешуй и располагаются мутовками в узлах. Здесь же образуются и боковые ветви. Ассимилирующую функцию выполняют зеленые стебли, поверхность которых увеличивается ребристостью, стенки клеток эпидермы пропитаны кремнеземом. Подземная часть представлена сильно развитым корневищем, в узлах которого формируются придаточные корни. У хвоща полевого (Equisetum arvense) боковые ветви корневища служат местом отложения запасных веществ, а также органами вегетативного размножения (рис. 11. 6).

Рис. 11. 6. Хвощ полевой: а, б – вегетативный и спороносный побеги спорофита; в – спорангиофор со спорангиями; г, д – споры; е – мужской гаметофит с антеридиями; ж – сперматозоид; з – обоеполый гаметофит; и — архегонии.
Весной на обычных или специальных спороносных стеблях образуются колоски, состоящие из оси, которая несет особые структуры, имеющие вид шестигранных щитков (спорангиофоры). Последние несут 6-8 спорангиев. Внутри спорангиев образуются споры, одетые толстой оболочкой, снабженной гигроскопичными лентовидными выростами – элатерами. Благодаря элатерам споры сцепляются вместе в комки, хлопья. Групповое распространение спор способствует тому, что при их прорастании разнополые заростки оказываются рядом, и это облегчает оплодотворение.
Заростки имеют вид небольшой длинно-лопастной зеленой пластинки с ризоидами на нижней поверхности. Мужские заростки меньше женских и несут по краям лопастей антеридии с многожгутиковыми сперматозоидами. На женских заростках в средней части развиваются архегонии. Оплодотворение происходит при наличии воды. Из зиготы развивается зародыш нового растения – спорофита.
В настоящее время большой роли в образовании растительного покрова хвощи не имеют. В лесах, на избыточно увлажненной почве, широко распространен хвощ лесной (E. sylvaticum) с сильно ветвящимися, поникающими боковыми веточками. На лугах, паровых полях, в посевах встречается трудно искореняемый сорняк хвощ полевой (E. arvense). У этого хвоща рано весной появляются неветвящиеся побеги, несущие спороносные колоски. Позднее от корневища развиваются зеленые вегетативные побеги. В лесной зоне на песчаных почвах и в оврагах широко распространен хвощ зимующий (E. hyemale).
Вегетативные побеги хвоща полевого (E. arvense) в официнальной медицине применяют: в качестве мочегонного средства при отеках на почве сердечной недостаточности; при заболеваниях мочевого пузыря и мочевыводящих путей; как кровоостанавливающее средство при маточных кровотечениях; при некоторых формах туберкулеза.
Отдел Папоротниковидные – Polypodiophyta
Возникли папоротники в девоне, когда древовидные папоротники вместе с ныне ископаемыми плаунами и хвощами господствовали в растительном покрове земли. Большинство их вымерло, оставшиеся дали начало мезозойским формам, которые были представлены очень широко. Папоротники намного превосходят по числу современных видов все остальные отделы высших споровых (около 25000).
У большинства живущих сейчас (исключая тропические) папоротников отсутствует наземный прямостоячий стебель, но есть подземный в виде корневища. От корневища отходят придаточные корни и крупные листья (вайи), имеющие стеблевое происхождение и длительно нарастающие верхушкой. Молодые листья обычно свернуты «улиткой». Среди существующих в настоящее время папоротников имеются как равноспоровые, так и разноспоровые.
В лесах Австралии, Южной Америки, Азии произрастают древовидные представители с колонновидными, неветвящимися, до 20 метров высоты стволами. В средней полосе нашей страны папоротники – это многолетние корневищные травы. Многие папоротники, как и мхи, являются индикаторами почв и типов леса. В светлых лесах, на песчаных или сухих подзолистых почвах распространен орляк обыкновенный (Pteridium aquilinum); на влажных богатых почвах кочедыжники (Athyrium) и крупные лесные щитовники (Dryopteris)(рис. 11. 7).
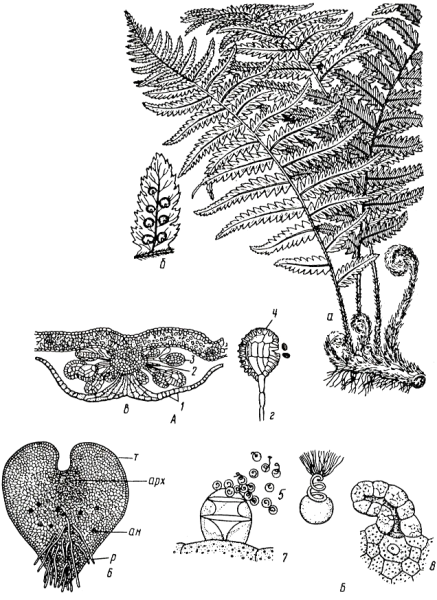
Рис. 11. 7. Щитовник мужской: А – спорофит: а – общий вид; б – сорусы на нижней стороне вайи; в – разрез соруса (1 – индузий, 2 – планцента, 3 – спорангий); г – спорангий (4 – кольцо); Б – гаметофит: 5 – сперматозоиды; 6 – заросток с нижней стороны (т – таллом, р – ризоиды, арх — архегонии, ан – антеридии); 7 – выход сперматозоидов из антеридия; 8 – архегоний с яйцеклеткой.
Цикл развития равноспоровых папоротников
В середине лета на нижней стороне зеленых листьев (у некоторых на специальных спороносных листьях) в виде коричневых бородавочек появляются группы спорангиев (сорусы). Сорусы у многих папоротников прикрыты сверху своеобразным покрывалом — индузием. Спорангии образуются на особом выросте листа (плаценте) и имеют чечевицеобразную форму, длинные ножки и многоклеточные стенки. В спорангиях хорошо выражено механическое кольцо, которое имеет вид узкой несмыкающейся полосы, опоясывающей спорангий. Когда кольцо подсыхает, стенки спорангия разрываются, и споры высыпаются.
Споры, образовавшиеся в спорангиях, одноклеточны и имеют толстую оболочку. При созревании они разносятся током воздуха и в благоприятных условиях прорастают, образуя сердцевидную зеленую многоклеточную пластинку (заросток), прикрепляющуюся к почве ризоидами. Заросток представляет собой половое поколение папоротников (гаметофит). На нижней стороне заростка образуются антеридии (со сперматозоидами) и архегонии (с яйцеклетками). При наличии воды сперматозоиды проникают в архегонии и оплодотворяют яйцеклетки. Из зиготы развивается зародыш, имеющий все основные органы (корень, стебель, лист и особый орган – ножку, прикрепляющую его к заростку). Постепенно зародыш начинает существовать самостоятельно, а заросток отмирает.
У разноспоровых папоротников гаметофиты редуцированы до микроскопических размеров (особенно мужские).
Из корневищ папоротника мужского (Dryopteris filix-mas), получают густой экстракт, который является эффективным противоглистным средством (ленточные черви).