Лист
Ботаника. Учебное пособие по специальности «Фармация»
Лист – боковая структурная часть побега, выполняющая функции фотосинтеза, газообмена и транспирации. Первые листовые органы семенных растений – семядоли зародыша. Все последующие листья возникают экзогенно на апексе побега в виде бугорков или выступов – листовых зачатков, которые впоследствии развиваются в листья (рис. 4.17).
Листья, как правило, имеют плоскую форму и дорсовентральное (спинно-брюшное) строение, в отличие от цилиндрических и радиально-симметричных осевых органов – стебля и корня. У семенных растений листья имеют ограниченный рост, так как, в отличие от осевых органов, лист растет за счет вставочных меристем и не имеет собственного меристематического апекса. Достигнув определенных размеров, лист до конца жизни остается без изменений.
Лист, как правило, не производит никаких других органов. В редких случаях на листе могут образоваться придаточные почки и придаточные корни (бегонии, бриофиллум), но непосредственно на листе новый лист обычно не образуется. Сам же лист всегда располагается только на оси побега – стебле.
Плоская пластинчатая форма листа создает наибольшую поверхность на единицу объема тканей, что способствует лучшему выполнению основной функции типичного зеленого листа – фотосинтеза. Плоская форма делает лист двусторонним. В зависимости от ориентации по отношению к верхушке побега выделяют верхнюю и нижнюю стороны листа. Дорсовентральность листа заключается в том, что в типичном случае верхняя и нижняя стороны резко отличаются по анатомическому строению, по характеру жилок, опушению и даже по окраске. На нижней стороне жилки более выпуклые, лист опушен сильнее, окраска нижней стороны листа более бледная, а иногда имеет фиолетовый или красный оттенок за счет антоцианов.
Морфология листа. Основной частью листа является листовая пластинка. Нижнюю часть листа, сочлененную со стеблем, называют основанием листа. Довольно часто между основанием и пластинкой формируется стеблеподобный цилиндрический или полукруглый в сечении черешок листа (рис. 4.51, 1). В этом случае листья называют черешковыми, в отличие от сидячих листьев, не имеющих черешка (рис. 4.51, 2). Роль черешка, кроме опорной и проводящей, состоит в том, что он долго сохраняет способность к вставочному росту и может регулировать положение пластинки, изгибаясь по направлению к свету.
Основание листа может принимать различную форму. Иногда оно почти незаметно или имеет вид небольшого утолщения (листовая подушечка), например у кислицы (рис. 4.51, 3). Часто основание разрастается, охватывая целиком узел и образуя трубку, называемую влагалищем листа (рис. 4.51, 4). Образование влагалища особенно характерно для однодольных, в частности для злаков, а из двудольных – для зонтичных. Влагалища защищают вставочные меристемы, находящиеся в основании междоузлий, и пазушные почки, сидящие над узлами.

Рис. 3.51. Части листа (схема): 1 – черешковый лист; 2 – сидячий лист; 3 – лист с подушечкой в основании; 4 – влагалищные листья; 5 – лист со свободными прилистниками; 6 – лист с приросшими к черешку прилистниками; 7 – лист с пазушными прилистниками; Пл – пластинка; Ос – основание; Вл – влагалище; Пр – прилистники; Ч – черешок; ПП – пазушная почка; ИМ – интеркалярная (вставочная) меристема.
Часто основание листа дает парные боковые выросты – прилистники. Форма и размеры прилистников различны у разных растений (рис. 4.51, 5,6,7). У древесных растений прилистники обычно имеют вид пленчатых чешуевидных образований и играют защитную роль, составляя главную часть почечных покровов. При этом они недолговечны и опадают при развертывании почек, так что на взрослом побеге у вполне развитых листьев прилистники не обнаруживаются (береза, дуб, липа, черемуха). Иногда прилистники имеют зеленую окраску и функционируют наравне с пластинкой листа как фотосинтезирующие органы (многие бобовые и розоцветные).
Для всех представителей семейства гречишных характерно образование раструбов. Раструб образуется в результате слияния двух пазушных прилистников и охватывает стебель над узлом в виде короткой пленчатой трубки.
Главная часть ассимилирующего листа — его пластинка. Если у листа одна пластинка, его называют простым. У сложных листьев на одном черешке с общим основанием располагаются две, три или несколько обособленных пластинок, иногда с собственными черешочками. Отдельные пластинки носят название листочков сложного листа, а общую ось, несущую листочки, называют рахисом. В зависимости от расположения листочков на рахисе различают перисто— и пальчатосложные листья. У первых листочки располагаются двумя рядами по обе стороны рахиса, продолжающего черешок. У пальчатосложных листьев рахиса нет, и листочки отходят от верхушки черешка. Частный случай сложного листа – тройчатосложный (рис. 4.52; 4.53).
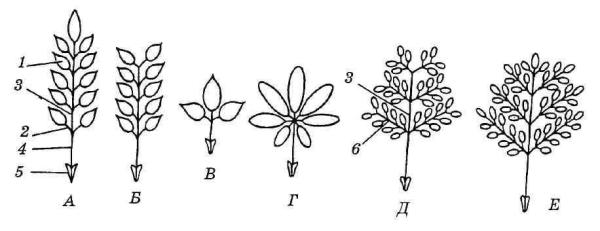
Рис. 4.52. Сложные листья (схема): А – непарноперистосложный; Б – парноперистосложный; В – тройчатосложный; Г – пальчатосложный; Д – дважды парноперистосложный; Е – дважды непарноперистосложный; 1 – листочек; 2 – черешочек; 3 – рахис; 4 – черешок; 5 – прилистники; 6 – рахис второго порядка.
Процесс формирования сложного листа напоминает ветвление, которое может идти до второго-третьего порядка, и тогда образуются дважды и трижды перистосложные листья. Если рахис заканчивается непарным листочком, лист называется непарноперистосложным, если парой листочков – парноперистосложным (рис. 4.52).
Форма пластинок простых листьев и листочков сложных листьев очень разнообразна. По форме листьев можно различать виды и роды растений в природе.
Пластинка листа или листочка может быть цельной или расчлененной более или менее глубоко на лопасти, доли или сегменты, располагающиеся при этом перисто или пальчато. Различают перисто— и пальчатолопастные, перисто— и пальчатораздельные и перисто— и пальчаторассеченные листья (рис. 4.53). Встречаются дважды, трижды и многократно расчлененные листовые пластинки.

Рис. 4.53. Типы расчленения пластинки простого листа.
Формы цельных листовых пластинок и расчлененных листьев в общем очертании выделяют в зависимости от двух параметров: соотношения между длиной и шириной и того, в какой части пластинки находится ее наибольшая ширина (рис. 4.54).
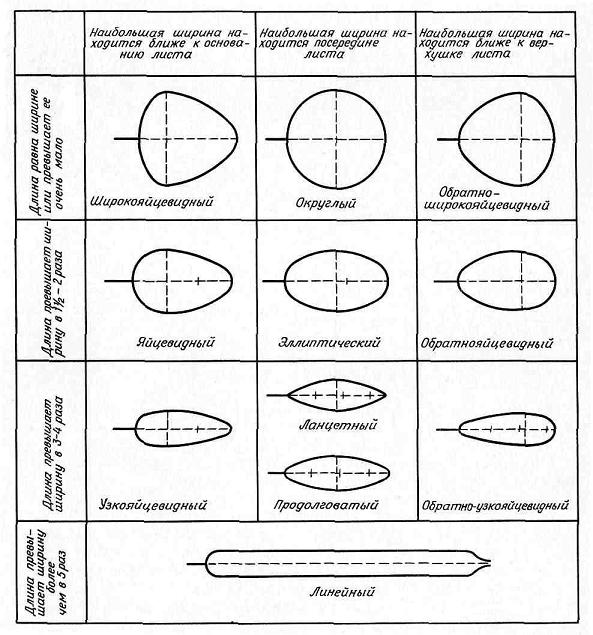
Рис. 4.54. Обобщенная схема форм листьев.
Некоторые особые формы листовых пластинок приведены на рис. 4.55.
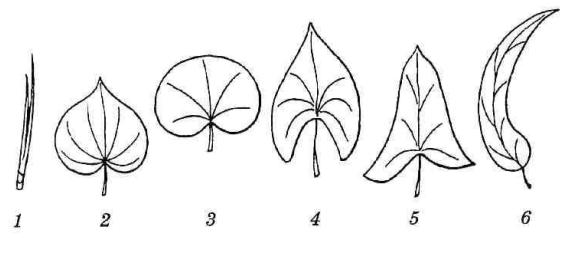
Рис. 4.55. Формы пластинок листьев: 1 – игольчатая; 2 – сердцевидная; 3 – почковидная; 4 – стреловидная; 5 – копьевидная; 6 – серповидная.
При описании также обращают внимание на форму верхушки, основания и края пластинки (рис. 4.56).
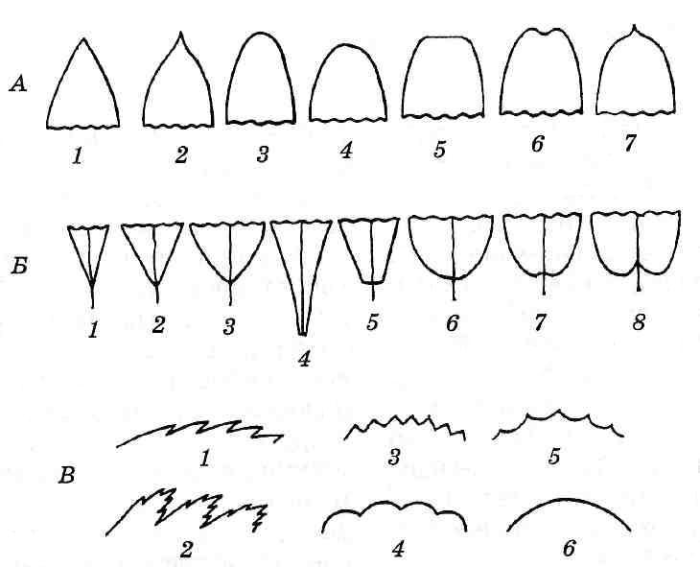
Рис. 4.56. Основные типы верхушек, оснований и края листовых пластинок: А – верхушки: 1 – острая; 2 – заостренная; 3 – тупая; 4 – округлая; 5 – усеченная; 6 — выемчатая; 7 – остроконечная; Б – основания: 1 – узкоклиновидное; 2 – клиновидное; 3 – ширококлиновидное; 4 – низбегающее; 5 – усеченное; 6 – округлое; 7 – выемчатое; 8 – сердцевидное; В – край листа: 1 – пильчатый; 2 – двоякопильчатый; 3 — зубчатый; 4 – городчатый; 5 – выемчатый; 6 – цельный.
Один из важных описательных признаков листа — характер жилкования (рис. 4.57). Жилкование – это система проводящих пучков и сопровождающих их тканей, посредством которых осуществляется транспорт веществ в листе. Наиболее примитивным является дихотомическое, или вильчатое жилкование, при котором жилки первого порядка на верхушке разделяются на две жилки второго порядка и т. д. (папоротники, гинкго двулопастный). У большинства хвойных в листе проходит одна или несколько жилок, не связанных между собой (простое жилкование).
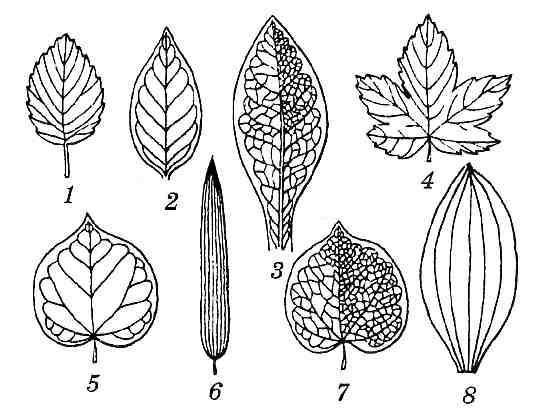
Рис. 4.57. Основные типы жилкования листьев покрытосеменных растений: 1 – перистокраевое; 2 – перистопетлевидное; 3 – перистосетчатое; 4 – пальчатокраевое; 5 – пальчатопетлевидное; 6 – параллельное; 7 – пальчатосетчатое; 8 – дуговидное.
У однодольных жилки проходят вдоль листа, не сливаясь друг с другом или отчасти сливаясь близ верхушки. Между собой они соединяются сетью мелких поперечных жилок – аностомозов. В зависимости от особенностей прохождения жилок выделяют параллельное и дуговидное жилкования (рис. 4.57). При этом лист называют параллельнонервным и дугонервным.
У двудольных растений имеются два основных типа жилкования – перистое и пальчатое. У листьев с перистым жилкованием (перистонервный лист) имеется одна главная жилка, являющаяся продолжением черешка. Она проходит от основания пластинки к ее верхушке. От главной жилки под углом отходят боковые жилки первого порядка, от них жилки второго порядка и т. д. Пальчатонервный лист не имеет главной жилки. У листьев с пальчатым жилкованием от места сочленения черешка и пластинки отходят несколько пальцевидно расходящихся крупных жилок первого порядка. Если жилки первого порядка доходят до края пластинки, выделяют перисто— и пальчатокраевое (краебежное) жилкования. Если боковые жилки образуют петли, сливаясь не доходя до края листа, выделяют перисто— и пальчатопетлевидное жилкования. Наконец, часто боковые жилки обильно ветвятся, образуя густую сеть аностомозирующих мелких жилок — перисто— и пальчатосетчатое жилкования (рис. 4.57). Мелкие жилки образуют замкнутые участки – ареолы.
Размеры, форма и степень рассеченности листьев, хотя и являются наследственными признаками вида, очень изменчивы и зависят также от условий обитания особей. Очень разнообразно опушение листьев. Растения засушливых местообитаний имеют более обильное опушение, чем растения, живущие в условиях влажного климата. Считается, что густой покров волосков удерживает молекулы водяного пара и тем самым снижает интенсивность транспирации.
Размеры листьев чаще всего колеблются в пределах от 3 до15 см. Самые крупные листья характерны для растений влажных тропических лесов, живущих в наиболее благоприятных условиях (древовидные папоротники, пальмы, бананы, дынное дерево). Очень крупными являются плавающие листья некоторых водных растений: кувшинок, лотосов. Самые крупные – листья амазонской кувшинки виктории королевской диаметром до 2 м.
Во флоре лесной умеренной зоны Европы относительно наиболее крупные листья у некоторых деревьев и кустарников лиственных лесов, которые так и называют широколиственными (дуб, липа, клен, ясень, бук, орех), а также у лесных трав, которые именуют широкотравьем (медуница, копытень, сныть). У этих крупнолистных растений преобладают листья цельные или слабо расчлененные, по очертанию они округлые, почковидные, широкояйцевидные, часто с сердцевидным основанием. В условиях более сухого и холодного климата Западной Сибири произрастают так называемые мелколиственные древесные растения с более мелкими листьями (береза, тополь, ольха, осина). Листья растений, выросших в условиях прямого и сильного освещения, обычно гораздо более мелкие и узкие по сравнению с родственными видами более тенистых и влажных местообитаний. Во флоре лугов и степей преобладают травы с листьями линейными, ланцетными, а также сильно рассеченными.
Анатомическое строение листа. Особенности строения листа определяются его главной функцией – фотосинтезом. Поэтому важнейшей частью листа является мезофилл, в котором сосредоточены хлоропласты, и происходит фотосинтез. Остальные ткани обеспечивают нормальную работу мезофилла. Эпидерма, покрывающая лист, регулирует газообмен и транспирацию. Система разветвленных проводящих пучков снабжает лист водой, необходимой для нормального протекания фотосинтеза, и обеспечивает отток ассимилятов. Наконец, механические ткани обеспечивают прочность листа.
Из всех органов лист в наибольшей степени связан с окружающей средой. Поэтому его строение гораздо сильнее, чем строение стебля или корня, отражает влияние изменчивых условий среды. Внешнее морфологическое разнообразие листьев сопровождается таким же разнообразием их анатомического строения.
Мезофилл занимает все пространство между верхней и нижней эпидермой, исключая проводящие и механические ткани. Клетки мезофилла довольно однородны, по форме чаще всего округлые или слегка вытянутые. Клеточные стенки остаются тонкими и неодревесневшими. Протопласт состоит из постенного слоя цитоплазмы с ядром и многочисленными хлоропластами. В центре клетки находится крупная вакуоль. Иногда стенки клеток образуют складки, которые увеличивают поверхность постенного слоя цитоплазмы и позволяют разместить большее число хлоропластов.
У большинства растений мезофилл дифференцирован на палисадную (столбчатую) и губчатую ткани (рис. 4.58).
Клетки палисадного мезофилла, расположенного, как правило, под верхней эпидермой, вытянуты перпендикулярно поверхности листа и образуют один или несколько слоев. Клетки губчатого мезофилла соединены более рыхло, межклетные пространства здесь могут быть очень большими по сравнению с объемом самих клеток. Увеличение межклетных пространств часто достигается тем, что клетки губчатого мезофилла образуют выросты.
Палисадная ткань содержит примерно три четверти всех хлоропластов листа и выполняет главную работу по ассимиляции углекислого газа. Поэтому палисадная ткань располагается в наилучших условиях освещения, непосредственно под верхней эпидермой. Благодаря тому, что клетки вытянуты перпендикулярно поверхности листа, лучи света легче проникают вглубь мезофилла.
Через губчатый мезофилл происходит газообмен. Углекислый газ из атмосферы через устьица, расположенные главным образом в нижней эпидерме, проникает в большие межклетники губчатого мезофилла и свободно расходится внутри листа. Кислород, выделяемый при фотосинтезе, передвигается в обратном направлении и через устьица выходит в атмосферу. Расположение устьиц преимущественно на нижней стороне листа объясняется не только положением губчатого мезофилла. Потеря воды листом в процессе транспирации идет медленнее через устьица, расположенные в нижней эпидерме. Кроме того, главным источником углекислого газа в атмосфере является «почвенное дыхание», т. е. выделение СО2 в результате дыхания многочисленных живых существ, населяющих почву.
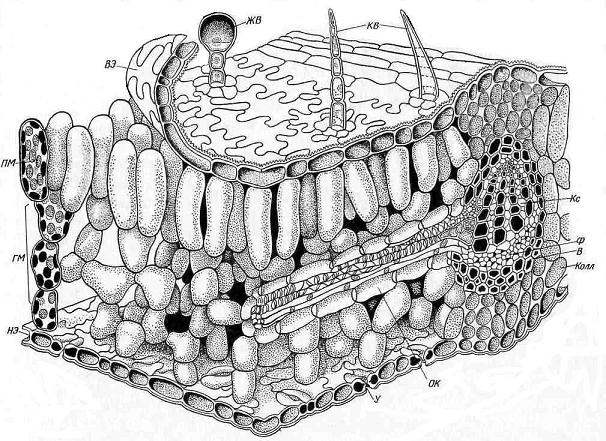
Рис. 4.58. Объемное изображение части листовой пластинки: В – волокна; ВЭ – верхняя эпидерма; ГМ – губчатый мезофилл; ЖВ – железистый волосок; КВ – кроющий волосок; Колл – колленхима; Кс – ксилема; НЭ – нижняя эпидерма; ОК – обкладочные клетки пучка; ПМ – палисадный мезофилл; У – устьице; Ф – флоэма.
Толщина палисадной и губчатой ткани и число слоев клеток в них различны в зависимости от условий освещения. Даже в пределах одной особи листья, выросшие на свету (рис. 4.59), имеют более развитый столбчатый мезофилл, чем листья, выросшие в условиях затенения (рис. 4.60).
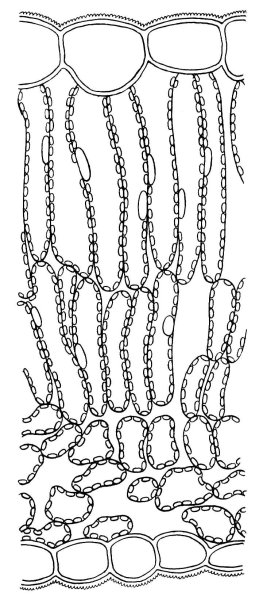
Рис. 4.59. Поперечный срез светового листа сирени.
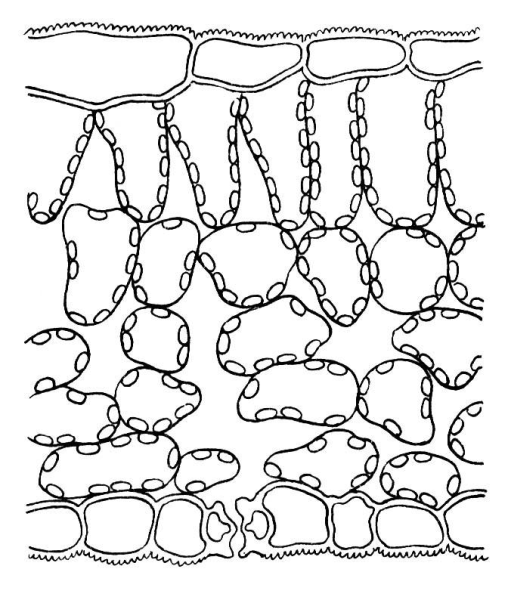
Рис. 4.60. Поперечный срез теневого листа сирени.
У тенелюбивых лесных растений палисадный мезофилл состоит из одного слоя клеток, имеющих характерную форму широко раскрытых воронок (рис. 4.61). Крупные хлоропласты расположены в них так, что не затеняют друг друга. Губчатый мезофилл также состоит из одного – двух слоев. Наоборот, у растений открытых местообитаний палисадный мезофилл насчитывает несколько слоев клеток и имеет значительную общую толщину (рис. 4.62).
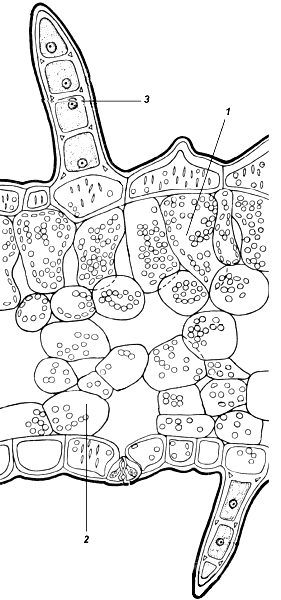
Рис. 4.61. Поперечный срез листа копытня: 1 – столбчатый мезофилл; 2 – губчатый мезофилл; 3 – кроющий волосок.
Листья, у которых палисадная ткань размещена на верхней стороне пластинки, а губчатая – на нижней, носят название дорсовентральных (рис. 4.58; 4.61; 4.62).
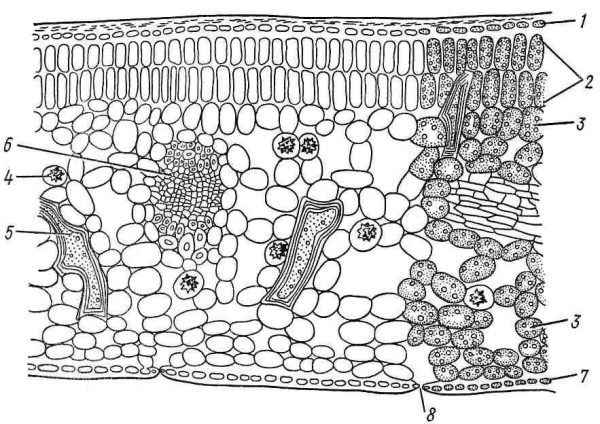
Рис. 4.62. Поперечный срез листа камелии: 1 – верхняя эпидерма; 2 – столбчатый мезофилл; 3 – губчатый мезофилл; 4 – клетка с друзой; 5 – склереида; 6 – проводящий пучок; 7 – нижняя эпидерма; 8 – устьице.
Если нижняя сторона листьев получает достаточно света, то и на ней образуется палисадный мезофилл (рис. 4.63). Листья с одинаковым мезофиллом на обеих сторонах называют изолатеральными.
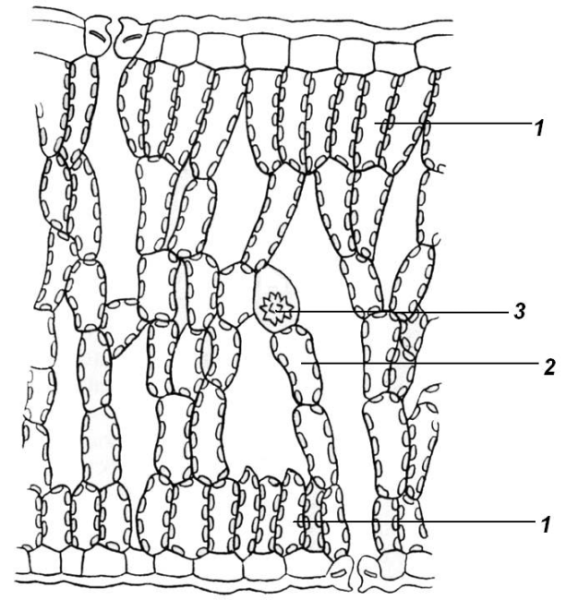
Рис. 4.63. Поперечный срез листа эвкалипта: 1 – столбчатый мезофилл; 2 – губчатый мезофилл; 3 – друза.
Не у всех растений мезофилл дифференцирован на палисадную и губчатую ткани, часто (особенно у однодольных растений) мезофилл совершенно однороден (рис. 4.64).
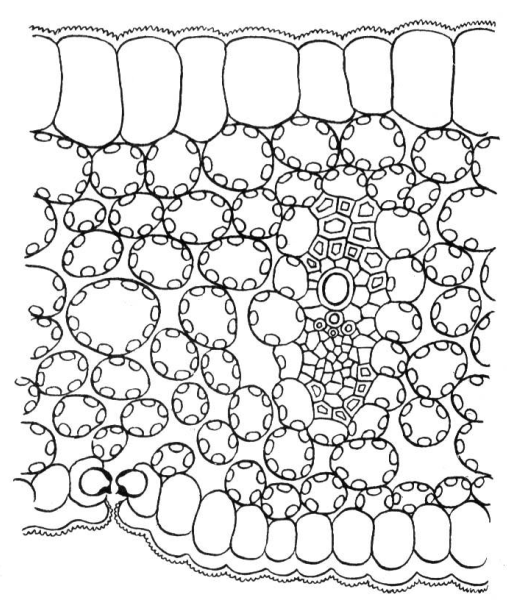
Рис. 4.64. Поперечный срез листа хлорофитума.
В мезофилле листьев часто встречаются клетки с кристаллами оксалата кальция, форма кристаллов играет большую роль в диагностике лекарственного растительного сырья.
Покровной тканью листа всегда является эпидерма. Вариации в ее строении зависят от условий обитания и выражаются в толщине кутикулы и восковых образований, в наличии разных типов трихомов, в характере, числе и размещении устьиц. На листьях, ориентированных верхней стороной к свету, устьица чаще размещены в нижней эпидерме (гипостоматические листья) (рис. 4.58; 4.62). При равномерном освещении обеих сторон устьица обычно имеются на обеих сторонах (амфистоматические листья) (рис. 4.63). Устьица могут располагаться исключительно на верхней стороне, например у листьев, плавающих на поверхности воды (эпистоматические листья) (рис. 3.65).
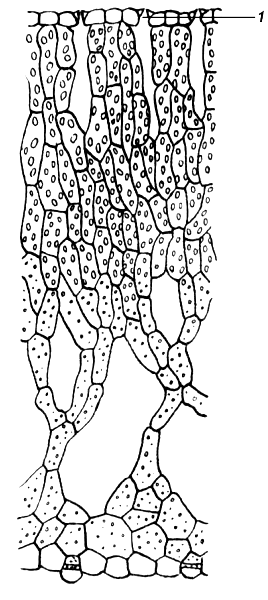
Рис. 4.65. Поперечный срез листа кубышки: 1 – устьице.
Проводящие ткани в листьях объединены в закрытые коллатеральные пучки (рис. 4.58; 4.66). Ксилема повернута к верхней, а флоэма – к нижней стороне листа. При такой организации проводящие ткани стебля и листьев образуют единую непрерывную систему.
Проводящие пучки с окружающими их тканями называют жилками. Крупные жилки часто сильно выдаются над поверхностью листа, особенно с нижней стороны. Менее крупные пучки полностью погружены в мезофилл. Жилки обычно образуют сеть с замкнутыми ячейками, однако самые мелкие из них могут иметь в мезофилле слепые окончания.
Проводящие элементы пучков непосредственно не соприкасаются с клетками мезофилла и межклетниками. В более крупных пучках они окружены склеренхимой, а в мелких – плотно сомкнутыми обкладочными клетками (рис. 4.58; 4.66). Обкладочные клетки отличаются от соседних клеток мезофилла более крупными размерами, они часто лишены хлоропластов. Обкладочные клетки аналогично эндодерме осевых органов регулируют ближний транспорт веществ в листе.
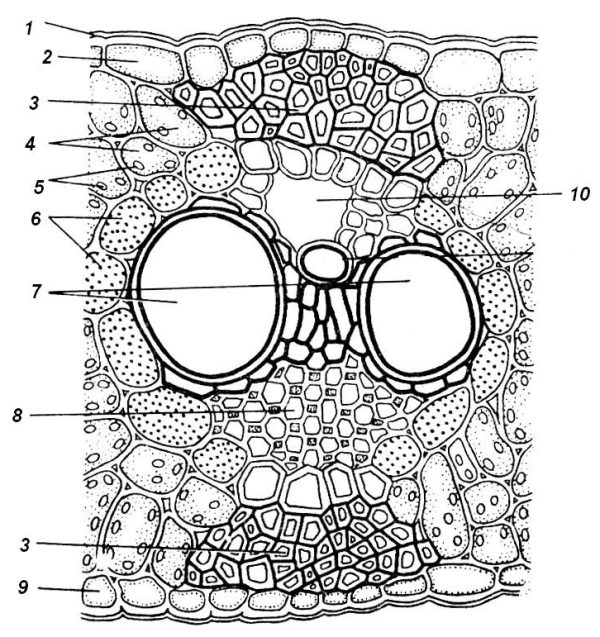
Рис. 4.66. Поперечный срез листа кукурузы в области крупного проводящего пучка: 1 – кутикула; 2 – верхняя эпидерма; 3 – склеренхима; 4 – клетки мезофилла; 5 – хлоропласты; 6 – обкладочные клетки; 7 – ксилема; 8 – флоэма; 9 – нижняя эпидерма; 10 – воздушная полость.
Механические ткани листа играют роль арматуры и противостоят его разрыву и раздавливанию. Это склеренхимные волокна, отдельные склереиды и тяжи колленхимы. Сочетаясь с живыми упругими клетками мезофилла, механические элементы образуют нечто вроде железобетона. Надежно соединенные между собой, клетки эпидермы играют роль внешней обвязки, повышающей общую прочность листа.
Склеренхимные волокна чаще всего сопровождают крупные проводящие пучки. Они окружают проводящие ткани со всех сторон или только сверху и снизу (рис. 4.66). Колленхима чаще присутствует около крупных пучков или по краю листа, предохраняя его от разрыва (рис. 4.58). Склереиды различной формы встречаются в мезофилле некоторых видов растений, имеющих плотные кожистые листья (кубышка, камелия) (рис. 4.62).
Прочность листьев может быть очень высокой. У многих пальм листья достигают в длину нескольких метров, но, несмотря на ветер, ливневые дожди и пр., сохраняют форму и положение в пространстве.
Разнообразие листьев. Листья неодинаковы не только у разных растений, но и в пределах одного и того же растения. Первые листовые органы проростка – семядоли, как правило, и по форме, и по размерам отличаются от всех последующих листьев. Следующие за семядолями листья проростка и молодого растения образуют листовую серию, в которой иногда наблюдается лишь постепенное увеличение размеров листьев, а иногда – очень резкие изменения их формы в сторону усложнения.
В основании годичного побега нередко находятся чешуевидные пленчатые, бледно-зеленые, буроватые или с антоциановой окраской листья. Они не имеют листовых пластинок и представляют собой разросшиеся основания листа, выполняющие защитную функцию. Их называют листьями низовой формации. К этой категории листьев относятся почечные чешуи, семядоли, чешуи луковиц и др.
Зеленые ассимилирующие листья составляют срединную формацию. В области соцветия расположены мелкие, пленчатые, зеленые или окрашенные в другой цвет листья. Это листья верховой формации, они служат кроющими листьями отдельных цветков или веточек соцветия, защищая их в почке. Они мельче и проще по очертаниям, чем срединные. Иногда их редукция может доходить до формирования мелких незеленых пленчатых чешуек – специализированных прицветников.
Разнообразие форм листьев на одном и том же растении в пределах срединной формации носит название гетерофиллии (разнолистности). Подобные различия могут быть связаны не только с возрастными изменениями, но и с влиянием внешних условий. Это особенно хорошо выражено у водных растений, побеги которых имеют погруженные и надводные части, например стрелолист, поручейник, водяной лютик (рис. 4.67). Подводные листья этих растений, лентовидные или многократно нитевидно рассеченные, отличаются от надводных – цельных или лопастных.
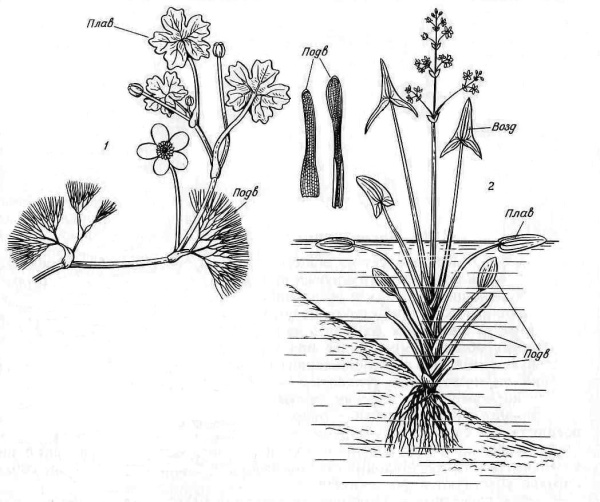
Рис. 4.67. Гетерофиллия у водных растений: 1 – водяной лютик; 2 – стрелолист; Подв – подводные листья; Плав – плавающие листья; Возд – воздушные листья.
Анизофиллией называют различия в форме и размерах ассимилирующих листьев на одном и том же узле побега (при супротивном или мутовчатом листорасположении). Чаще всего анизофиллия наблюдается у плагиотропных побегов древесных и травянистых растений. Разница размеров обусловлена действием силы тяжести и различием в освещенности верхней и нижней стороны побега.
Длительность жизни листьев намного меньше, чем длительность жизни осевых органов. Это связано с особенностями их функционирования в качестве органов фотосинтеза. Чрезвычайно высокая метаболическая активность приводит к быстрому старению и отмиранию тканей листа.
У большинства растений продолжительность жизни листьев не превышает одного-полутора астрономических лет (чаще – 4-5 месяцев). От 2 до 5 лет живут листья у ряда субтропических растений, а также у растений тайги, тундры и высокогорий. Наибольшей продолжительностью жизни отличаются листья хвойных – до 15 лет и более.
Многолетние растения, которые круглый год несут зеленые листья, называют вечнозелеными, в отличие от листопадных, пребывающих хотя бы недолго в безлистном состоянии. Вечнозеленые деревья, кустарники и кустарнички характерны для влажных тропических и субтропических лесов, для хвойных лесов умеренной зоны и для различных типов тундровой растительности.
У листопадных деревьев и кустарников опадение листьев на зиму имеет важное приспособительное значение. Наибольшую опасность зимой представляет высыхание надземных органов растений, так как потеря влаги в это время года не может быть компенсирована. Сбрасывая листья, растения резко уменьшают испаряющую поверхность; остающиеся органы – ствол и ветви – надежно защищены вторичными покровными тканями. Опасность заключается и в возможности поломок облиственных ветвей от тяжести снега, тогда как на безлистных ветвях снег не накапливается. Для древесных растений, живущих в безморозном климате с резко выраженным засушливым периодом, листопад также представляет приспособление к перенесению засухи.
По мере старения листа постепенно снижается интенсивность фотосинтеза и дыхания, а также содержание белков и РНК. Видимый признак старения листа – его пожелтение или покраснение, связанное с деградацией хлоропластов, разрушением хлорофилла и накоплением каротиноидов и антоцианов. В тканях старых листьев откладываются в большом количестве кристаллы оксалата кальция. Из листьев оттекают пластические вещества; лист перед опадением «опустошается».
У двудольных древесных растений близ основания листа формируется отделительный слой, состоящий из легко расслаивающейся паренхимы. По этому слою лист отделяется от стебля, причем на поверхности будущего листового рубца заранее образуется защитный слой пробки. У однодольных и травянистых двудольных отделительного слоя не образуется, лист отмирает и разрушается постепенно, оставаясь на стебле.
У вечнозеленых растений массовый листопад чаще всего приурочен к началу роста новых побегов из почек. В частности, у хвойных растений и вечнозеленых трав массовое отмирание и опадение листьев наблюдается не осенью, а весной.