Корень и корневые системы
Ботаника. Учебное пособие по специальности «Фармация»
Корень – основной вегетативный орган растения, выполняющий в типичном случае функцию почвенного питания. Корень – осевой орган, обладающий радиальной симметрией и неопределенно долго нарастающий в длину благодаря деятельности апикальной меристемы. От побега он морфологически отличается тем, что на нем никогда не образуются листья, а апикальная меристема всегда прикрыта корневым чехликом.
Кроме главной функции поглощения веществ из почвы, корни выполняют и другие функции:
- корни укрепляют («заякоривают») растения в почве, делают возможным вертикальный рост и вынесение побегов наверх;
- в корнях синтезируются различные вещества, которые затем передвигаются в другие органы растения;
- в корнях могут откладываться запасные вещества;
- корни взаимодействуют с корнями других растений, микроорганизмами, грибами, обитающими в почве.
Совокупность корней одной особи образует единую в морфологическом и физиологическом отношении корневую систему.
В состав корневых систем входят корни различной морфологической природы – главный корень, боковые и придаточные корни.
Главный корень развивается из зародышевого корешка. Боковые корни образуются на корне (главном, боковом, придаточном), который по отношению к ним обозначается как материнский. Они возникают на некотором расстоянии от апекса, в направлении от основания корня к его верхушке. Боковые корни закладываются эндогенно, т.е. во внутренних тканях материнского корня. Если бы ветвление происходило в самом апексе, это бы затруднило продвижение корня в почве. Придаточные корни могут возникать и на стеблях, и на листьях, и на корнях. В последнем случае они отличаются от боковых корней тем, что не обнаруживают строгого порядка заложения вблизи апекса материнского корня и могут возникать на старых участках корней.
По происхождению выделяют следующие типы корневых систем (рис. 4.1):
1) система главного корня представлена главным корнем (первого порядка) с боковыми корнями второго и последующих порядков (у многих кустарников и деревьев, большинства двудольных растений);
2) система придаточных корней развивается на стеблях, листьях; встречается у большинства однодольных растений и многих двудольных, размножающихся вегетативно;
3) смешанная корневая система образована главным и придаточными корнями с их боковыми ответвлениями (многие травянистые двудольные).

Рис. 4.1. Типы корневых систем: А – система главного корня; Б – система придаточных корней; В – смешанная корневая система (А и В – стержневые корневые системы; Б – мочковатая корневая система).
По форме различают стержневую и мочковатую корневые системы.
В стержневой корневой системе главный корень сильно развит и хорошо заметен среди остальных корней. В мочковатой корневой системе главный корень незаметен или его нет, а корневая система составлена многочисленными придаточными корнями (рис. 4.1).
Корень обладает потенциально неограниченным ростом. Однако в естественных условиях рост и ветвление корней ограничены влиянием других корней и почвенных экологических факторов. Основная масса корней располагается в верхнем слое почвы (15 см), наиболее богатом органическими веществами. Корни деревьев углубляются в среднем на 10-15 м, а в ширину распространяются обычно за пределы радиуса крон. Корневая система кукурузы заходит на глубину около 1,5 м и примерно на 1 м во все стороны от растения. Рекордная глубина проникновения корней в почву отмечена у пустынного мескитового кустарника – более 53 м.
У одного куста ржи, выращенного в теплице, общая длина всех корней составила 623 км. Суммарный прирост всех корней за одни сутки равнялся примерно 5 км. Общая поверхность всех корней у этого растения составила 237 м2 и была в 130 раз больше поверхности надземных органов.
Зоны молодого корневого окончания — это разные по длине части молодого корня, выполняющие неодинаковые функции и характеризующиеся определенными морфологическими и анатомическими особенностями (рис. 4.2).
Кончик корня снаружи всегда прикрыт корневым чехликом, защищающим апикальную меристему. Чехлик состоит из живых клеток и постоянно обновляется: по мере того, как с его поверхности слущиваются старые клетки, на смену им, изнутри, апикальная меристема образует новые молодые клетки. Наружные клетки корневого чехлика отслаиваются еще будучи живыми, они продуцируют обильную слизь, которая облегчает продвижение корня среди твердых частиц почвы. В клетках центральной части чехлика содержится много крахмальных зерен. По-видимому, эти зерна служат статолитами, т. е. способны перемещаться в клетке при изменении положения кончика корня в пространстве, благодаря чему корень растет всегда в сторону действия силы тяжести (положительный геотропизм).
Под чехликом находится зона деления, представленная апикальной меристемой, в результате деятельности которой формируются все прочие зоны и ткани корня. Зона деления имеет размеры около 1 мм. Клетки апикальной меристемы относительно мелкие, многогранные, с густой цитоплазмой и крупным ядром.
Вслед за зоной деления располагается зона растяжения, или зона роста. В этой зоне клетки почти не делятся, а сильно растягиваются (растут) в продольном направлении, вдоль оси корня. Объем клеток увеличивается за счет поглощения воды и образования крупных вакуолей, при этом высокое тургорное давление проталкивает растущий корень между частицами почвы. Протяженность зоны растяжения обычно невелика и не превышает нескольких миллиметров.

Рис. 4.2. Общий вид (А) и продольный срез (Б) корневого окончания (схема): I – корневой чехлик; II – зоны деления и растяжения; III – зона всасывания; IV – начало зоны проведения: 1 – растущий боковой корень; 2 – корневые волоски; 3 – ризодерма; 3а – экзодерма; 4 – первичная кора; 5 – эндодерма; 6 – перицикл; 7 – осевой цилиндр.
Далее идет зона поглощения, или зона всасывания. В этой зоне покровной тканью является ризодерма (эпиблема), клетки которой несут многочисленные корневые волоски. Растяжение корня прекращается, корневые волоски плотно охватывают частицы почвы и как бы срастаются с ними, поглощая воду и растворенные в ней минеральные соли. Зона поглощения имеет протяжение до нескольких сантиметров. Эту зону называют также зоной дифференциации, поскольку именно здесь происходит образование постоянных первичных тканей.
Продолжительность жизни корневого волоска не превышает 10-20 дней. Выше зоны всасывания, там, где исчезают корневые волоски, начинается зона проведения. По этой части корня вода и растворы солей, поглощенные корневыми волосками, транспортируются в вышележащие органы растения. В зоне проведения формируются боковые корни (рис. 4.2).
Клетки зон всасывания и проведения занимают фиксированное положение и не могут смещаться относительно участков почвы. Однако сами зоны, вследствие постоянного верхушечного роста, непрерывно перемещаются вдоль корня по мере нарастания корневого окончания. В зону поглощения постоянно включаются молодые клетки со стороны зоны растяжения и одновременно исключаются клетки стареющие, переходящие в состав зоны проведения. Таким образом, всасывающий аппарат корня – подвижное образование, непрерывно передвигающееся в почве.
Так же последовательно и закономерно в корневом окончании возникают внутренние ткани.
Первичное строение корня. Первичная структура корня образуется в результате деятельности апикальной меристемы. Корень отличается от побега тем, что его апикальная меристема откладывает клетки не только внутрь, но и наружу, пополняя чехлик. Число и расположение инициальных клеток в апексах корней значительно варьируют у растений, принадлежащих к разным систематическим группам. Производные инициалей уже вблизи апикальной меристемы дифференцируются в первичные меристемы – 1) протодерму, 2) основную меристему и 3) прокамбий (рис. 4.3). Из этих первичных меристем в зоне всасывания формируются три системы тканей: 1) ризодерма, 2) первичная кора и 3) осевой (центральный) цилиндр, или стела.

Рис. 4.3. Продольный срез кончика корня лука.
Ризодерма (эпиблема, эпидерма корня) – всасывающая ткань, образующаяся из протодермы, наружного слоя первичной меристемы корня. В функциональном отношении ризодерма представляет собой одну из важнейших тканей растения. Через нее идет поглощение воды и минеральных солей, она взаимодействует с живым населением почвы, через ризодерму из корня в почву выделяются вещества, помогающие почвенному питанию. Поглощающая поверхность ризодермы сильно увеличена благодаря наличию у части клеток трубчатых выростов – корневых волосков (рис. 4.4). Волоски имеют длину 1-2 мм (до 3 мм). У одного четырехмесячного растения ржи было установлено примерно 14 млрд. корневых волосков с площадью поглощения 401 м2 и суммарной длиной более 10 000 км. У водных растений корневые волоски могут отсутствовать.
Стенка волоска очень тонкая и состоит из целлюлозы и пектиновых веществ. Ее наружные слои содержат слизь, что способствует установлению более тесного контакта с частицами почвы. Слизь создает благоприятные условия для поселения полезных бактерий, влияет на доступность почвенных ионов и защищает корень от иссушения. В физиологическом отношении ризодерма отличается большой активностью. Она поглощает минеральные ионы с затратой энергии. В гиалоплазме имеется большое количество рибосом и митохондрий, что характерно для клеток с высоким уровнем обмена веществ.

Рис. 4.4. Поперечный срез корня в зоне всасывания: 1 – ризодерма; 2 – экзодерма; 3 – мезодерма; 4 — эндодерма; 5 – ксилема; 6 – флоэма; 7 — перицикл.
Из основной меристемы формируется первичная кора. Первичная кора корня дифференцирована на: 1) экзодерму – наружную часть, лежащую непосредственно за ризодермой, 2) среднюю часть – мезодерму и 3) самый внутренний слой – эндодерму (рис. 4.4). Основную массу первичной коры составляет мезодерма, образованная живыми паренхимными клетками с тонкими стенками. Клетки мезодермы расположены рыхло, по системе межклетников вдоль оси корня циркулируют газы, необходимые для дыхания клеток. У болотных и водных растений, корни которых испытывают недостаток кислорода, мезодерма часто представлена аэренхимой. Также в мезодерме могут присутствовать механические и выделительные ткани. Паренхима первичной коры выполняет ряд важных функций: участвует в поглощении и проведении веществ, синтезирует различные соединения, в клетках коры часто откладываются запасные питательные вещества, например крахмал.
Наружные слои первичной коры, подстилающие ризодерму, образуют экзодерму. Экзодерма возникает как ткань, регулирующая прохождение веществ из ризодермы в кору, но после отмирания ризодермы выше зоны всасывания она оказывается на поверхности корня и превращается в защитную покровную ткань. Экзодерма формируется как один слой (реже несколько слоев) и состоит из живых паренхимных клеток, плотно сомкнутых между собой. По мере отмирания корневых волосков, стенки клеток экзодермы покрываются с внутренней стороны слоем суберина. В этом отношении экзодерма сходна с пробкой, но в отличие от нее первична по происхождению, и клетки экзодермы остаются живыми. Иногда в экзодерме сохраняются пропускные клетки с тонкими неопробковевшими стенками, через которые происходит избирательное поглощение веществ.
Самый внутренний слой первичной коры – эндодерма. Она в виде непрерывного цилиндра окружает стелу. Эндодерма в своем развитии может пройти три ступени. На первой ступени ее клетки плотно прилегают друг к другу и имеют тонкие первичные стенки. На их радиальных и поперечных стенках образуются утолщения в виде рамочек – пояски Каспари (рис. 4.5). Пояски соседних клеток тесно смыкаются между собой, так что вокруг стелы создается их непрерывная система. В поясках Каспари откладываются суберин и лигнин, что делает их непроницаемыми для растворов. Поэтому вещества из коры в стелу и из стелы в кору могут пройти только по симпласту, т. е. через живые протопласты клеток эндодермы и под их контролем.

Рис. 4.5. Эндодерма на первой ступени развития (схема).
На второй ступени развития суберин откладывается по всей внутренней поверхности клеток эндодермы. При этом некоторые клетки сохраняют первичное строение. Это пропускные клетки, они остаются живыми, и через них осуществляется связь между первичной корой и центральным цилиндром. Как правило, они расположены напротив лучей первичной ксилемы. У корней, не обладающих вторичным утолщением, эндодерма может получить третичное строение. Оно характеризуется сильным утолщением и одревеснением всех стенок, или чаще сравнительно тонкими остаются стенки, обращенные наружу (рис. 4.7). Пропускные клетки сохраняются и в третичной эндодерме.
Центральный (осевой) цилиндр, или стела формируется в центре корня. Уже вплотную к зоне деления самый наружный слой стелы образует перицикл, клетки которого долго сохраняют характер меристемы и способность к новообразованиям. В молодом корне перицикл состоит из одного ряда живых паренхимных клеток с тонкими стенками (рис. 4.4). Перицикл выполняет несколько важных функций. У большинства семенных растений в нем закладываются боковые корни. У видов с вторичным ростом он участвует в формировании камбия и дает начало первому слою феллогена. В перицикле часто происходит образование новых клеток, входящих затем в его состав. У некоторых растений в перицикле возникают также зачатки придаточных почек. В старых корнях однодольных растений клетки перицикла часто склерифицируются.
За перициклом находятся клетки прокамбия, которые дифференцируются в первичные проводящие ткани. Элементы флоэмы и ксилемы закладываются по кругу, чередуясь друг с другом, и развиваются центростремительно. Однако ксилема в своем развитии обычно обгоняет флоэму и занимает центр корня. На поперечном разрезе первичная ксилема образует звезду, между лучами которой располагаются участки флоэмы (рис. 4.4). Такая структура получила название радиального проводящего пучка.
Звезда ксилемы может иметь различное число лучей – от двух до многих. Если их два, корень называют диархным, если три – триархным, четыре – тетрархным, а если много – полиархным (рис. 4.6). Число лучей ксилемы обычно зависит от толщины корня. В толстых корнях однодольных растений оно может достигать 20-30 (рис. 4.7). В корнях одного и того же растения число лучей ксилемы может быть различно, в более тонких ответвлениях оно сокращается до двух.

Рис. 4.6. Типы строения осевого цилиндра корня (схема): А – диархный; Б – триархный; В – тетрархный; Г – полиархный: 1 – ксилема; 2 – флоэма.
Пространственное разделение тяжей первичной флоэмы и ксилемы, расположенных на разных радиусах, и их центростремительное заложение представляют собой характерные особенности строения центрального цилиндра корня и имеют большое биологическое значение. Элементы ксилемы максимально приближены к поверхности стелы, и в них легче, минуя флоэму, проникают растворы, поступающие из коры.

Рис. 4.7. Поперечный срез корня однодольного растения: 1 – остатки ризодермы; 2 – экзодерма; 3 – мезодерма; 4 – эндодерма; 5 – пропускные клетки; 6 – перицикл; 7 – ксилема; 8 – флоэма.
Центральная часть корня обычно занята одним или несколькими крупными сосудами ксилемы. Присутствие сердцевины вообще нетипично для корня, однако в корнях некоторых однодольных в середине находится небольшой участок механической ткани (рис. 4.7) или тонкостенные клетки, возникающие из прокамбия (рис. 4.8).

Рис. 4.8. Поперечный срез корня кукурузы.
Первичная структура корня характерна для молодых корней всех групп растений. У споровых и однодольных растений первичное строение корня сохраняется в течение всей жизни.
Вторичное строение корня. У голосеменных и двудольных растений первичное строение сохраняется недолго и выше зоны всасывания сменяется вторичным. Вторичное утолщение корня происходит за счет деятельности вторичных боковых меристем – камбия и феллогена.
Камбий возникает в корнях из меристематических прокамбиальных клеток в виде прослойки между первичными ксилемой и флоэмой (рис. 4.9). В зависимости от числа флоэмных тяжей одновременно закладываются две или более зоны камбиальной активности. Сначала камбиальные прослойки разобщены между собой, но вскоре клетки перицикла, лежащие против лучей ксилемы, делятся тангенциально и соединяют камбий в непрерывный слой, окружающий первичную ксилему. Камбий откладывает внутрь слои вторичной ксилемы (древесины) и наружу вторичную флоэму (луб). Если этот процесс длится долго, то корни достигают значительной толщины.

Рис. 4.9. Заложение и начало деятельности камбия в корне проростка тыквы: 1 – первичная ксилема; 2 – вторичная ксилема; 3 – камбий; 4 – вторичная флоэма; 5 – первичная флоэма; 6 – перицикл; 7 – эндодерма.
Участки камбия, возникшие из перицикла, состоят из паренхимных клеток и не способны откладывать элементы проводящих тканей. Они образуют первичные сердцевинные лучи, представляющие собой широкие участки паренхимы между вторичными проводящими тканями (рис. 4.10). Вторичные сердцевинные, или лубодревесинные лучи возникают дополнительно при длительном утолщении корня, они обычно yже первичных. Сердцевинные лучи обеспечивают связь между ксилемой и флоэмой корня, по ним происходит радиальный транспорт различных соединений.
В результате деятельности камбия первичная флоэма оттесняется наружу и сдавливается. Звезда первичной ксилемы остается в центре корня, ее лучи могут сохраняться в течение длительного времени (рис. 4.10), но чаще центр корня заполняется вторичной ксилемой, и первичная ксилема становится незаметной.
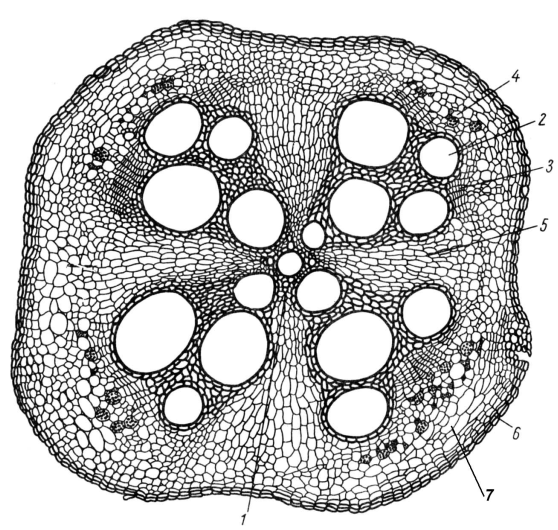
Рис. 4.10. Поперечный срез корня тыквы (вторичное строение): 1 – первичная ксилема; 2 – вторичная ксилема; 3 – камбий; 4 – вторичная флоэма; 5 – первичный сердцевинный луч; 6 – пробка; 7 – паренхима вторичной коры.
Ткани первичной коры не могут следовать за вторичным утолщением и обречены на гибель. Они заменяются вторичной покровной тканью – перидермой, которая может растягиваться на поверхности утолщающегося корня благодаря работе феллогена. Феллоген закладывается в перицикле и начинает откладывать наружу пробку, а внутрь — феллодерму. Первичная кора, отрезанная пробкой от внутренних живых тканей, отмирает и сбрасывается (рис. 4.11).
Клетки феллодермы и паренхима, образовавшаяся за счет деления клеток перицикла, образуют паренхиму вторичной коры, окружающую проводящие ткани (рис. 4.10). Снаружи корни вторичного строения покрыты перидермой. Корка образуется редко, лишь на старых корнях деревьев.
Многолетние корни древесных растений в результате длительной активности камбия нередко сильно утолщаются. Вторичная ксилема у таких корней сливается в сплошной цилиндр, окруженный снаружи кольцом камбия и сплошным кольцом вторичной флоэмы (рис. 4.11). По сравнению со стеблем границы годичных колец в древесине корня выражены значительно слабее, луб развит сильнее, сердцевинные лучи, как правило, шире.

Рис. 4.11. Поперечный срез корня ивы в конце первого вегетационного периода.
Специализация и метаморфозы корней. У большинства растений в одной и той же корневой системе отчетливо различаются ростовые и сосущие окончания. Ростовые окончания обычно более мощные, быстро удлиняются и продвигаются вглубь почвы. Зона растяжения у них хорошо выражена, и апикальные меристемы работают энергично. Сосущие окончания, возникающие в большом количестве на ростовых корнях, удлиняются медленно, и их апикальные меристемы почти перестают работать. Сосущие окончания как бы останавливаются в почве и интенсивно ее «обсасывают».
У древесных растений различают толстые скелетные и полускелетные корни, на которых образуются недолговечные корневые мочки. В состав корневых мочек, непрерывно заменяющих друг друга, входят ростовые и сосущие окончания.
Если корни выполняют особые функции, их строение меняется. Резкое, наследственно закрепленное видоизменение органа, вызванное сменой функций, носит название метаморфоза. Видоизменения корней очень разнообразны.
Корни многих растений образуют симбиоз с гифами почвенных грибов, называемый микоризой («грибокорень»). Микориза образуется на сосущих корнях в зоне поглощения. Грибной компонент облегчает корням получение воды и минеральных элементов из почвы, часто гифы грибов заменяют корневые волоски. В свою очередь, гриб получает от растения углеводы и другие питательные вещества. Различают два основных типа микоризы. Гифы эктотрофной микоризы образуют чехол, окутывающий корень снаружи. Эктомикориза широко распространена у деревьев и кустарников. Эндотрофная микориза встречается в основном у травянистых растений. Эндомикориза находится внутри корня, гифы внедряются в клетки коровой паренхимы. Микотрофное питание очень широко распространено. Некоторые растения, например орхидные, вообще не могут существовать без симбиоза с грибами.
На корнях бобовых возникают особые образования – клубеньки, в которых поселяются бактерии из рода Rhizobium. Эти микроорганизмы способны усваивать атмосферный молекулярный азот, переводя его в связанное состояние. Часть веществ, синтезированных в клубеньках, усваивают растения, бактерии, в свою очередь, используют вещества, находящиеся в корнях. Этот симбиоз имеет большое значение для сельского хозяйства. Бобовые растения благодаря дополнительному источнику азота богаты белками. Они дают ценные пищевые и кормовые продукты и обогащают почву азотистыми веществами.
Очень широко распространены запасающие корни. Они обычно утолщены и сильно паренхиматизированы. Сильно утолщенные придаточные корни называют корневыми шишками, или корнеклубнями (георгин, некоторые орхидные). У многих, чаще двулетних, растений со стержневой корневой системой возникает образование, носящее название корнеплода. В образовании корнеплода принимают участие и главный корень, и нижняя часть стебля. У моркови почти весь корнеплод составлен корнем, у репы корень образует лишь самую нижнюю часть корнеплода (рис. 4.12).

Рис.4.12. Корнеплоды моркови (1, 2), репы (3, 4) и свеклы (5, 6, 7) (на поперечных разрезах ксилема черная; горизонтальным пунктиром показана граница стебля и корня).
Корнеплоды культурных растений возникли в результате длительного отбора. В корнеплодах сильно развита запасающая паренхима и исчезли механические ткани. У моркови, петрушки и других зонтичных паренхима сильно развита во флоэме; у репы, редьки и других крестоцветных – в ксилеме. У свеклы запасные вещества откладываются в паренхиме, образованной деятельностью нескольких добавочных слоев камбия (рис. 4.12).
У многих луковичных и корневищных растений образуются втягивающие, или контрактильные корни (рис. 4.13, 1). Они могут укорачиваться и втягивать побег в почву на оптимальную глубину на время летней засухи или зимних морозов. Втягивающие корни имеют утолщенные основания с поперечной морщинистостью.

Рис. 4.13. Метаморфозы корня: 1 – клубнелуковица гладиолуса с утолщенными у основания втягивающими корнями; 2 – дыхательные корни с пневматофорами у авиценнии (пр – зона прилива); 3 – воздушные корни орхидеи.
Воздушные корни образуются у многих тропических эпифитных однодольных из семейств орхидных, ароидных, бромелиевых (рис. 4.13, 3). Эпифиты поселяются на других растениях, но не паразитируют на них, а используют как подпорку для поднятия вверх, к свету. Воздушные корни свободно висят в воздухе и приспособлены к поглощению атмосферной влаги. На их поверхности образуется веламен (рис. 4.14). Веламен, как и ризодерма, образуется из протодермы, но это многослойная ткань. Клетки веламена отмирают, их стенки имеют сетчатые или спиральные утолщения. Через поры и сквозные отверстия в клеточных стенках влага проникает капиллярным путем. Изнутри веламен подстилается экзодермой со сложно построенными пропускными клетками, через которые вода передается в клетки коры и осевого цилиндра (рис. 4.14). Веламен встречается и у некоторых наземных однодольных (кливия, хлорофитум), в этом случае он выполняет функцию механической защиты и предохраняет растение от потери воды из первичной коры.

Рис. 4.14. Часть поперечного среза воздушного корня орхидеи: 1 – веламен; 2 – экзодерма; 3 – пропускная клетка.
Дыхательные корни, или пневматофоры (рис. 4.13, 2) образуются у некоторых тропических древесных растений, живущих в условиях недостатка кислорода (таксодиум, или болотный кипарис; растения мангровых зарослей, обитающие по болотистым берегам океанических побережий). Пневматофоры растут вертикально вверх и высовываются над поверхностью почвы. Через систему отверстий в этих корнях, связанных с аэренхимой, воздух поступает в подводные органы.
У некоторых растений для поддержания побегов в воздушной среде образуются дополнительные опорные корни. Они отходят от горизонтальных ветвей кроны и, достигнув поверхности почвы, интенсивно ветвятся, превращаясь в столбовидные образования, поддерживающие крону дерева (столбовидные корни баньяна) (рис. 4.15, 2). Ходульные корни отходят от нижних участков стебля, придавая стеблю устойчивость. Они образуются у растений мангровых зарослей, растительных сообществ, развивающихся на затопляемых во время прилива тропических берегах океанов (рис. 4.15, 3), а также у кукурузы (рис. 4.15, 1). У фикуса каучуконосного образуются досковидные корни. В отличие от столбовидных и ходульных, они являются по происхождению не придаточными, а боковыми корнями.

Рис. 4.15. Опорные корни: 1 – ходульные корни кукурузы; 2 – столбовидные корни баньяна; 3 – ходульные корни ризофоры (пр – зона прилива; от – зона отлива; ил – поверхность илистого дна).
Побеги плюща, стремясь к солнцу, обвивают другие растения или прикрепляются к стенам при помощи корней — прицепок. У паразитических растений корни видоизменяются в присоски – гаустории, которые внедряются в ткани других растений и поглощают из клеток воду и питательные вещества