5.10. Клеточные структуры
Биология — Тейлор Д., Грин Н., Стаут У. 1 том — 2013
5.10.1. Ядро
Ядра имеются во всех эукариотических клетках, за исключением зрелых члеников ситовидных трубок флоэмы и зрелых эритроцитов млекопитающих. У некоторых протистов, в частности у Paramecium, имеется два ядра — микронуклеус и макронуклеус. Однако, как правило, клетки содержат только одно ядро. При рассмотрении клеток с помощью светового микроскопа ядра сразу бросаются в глаза, потому что из всех клеточных органелл они самые крупные. По этой же причине именно они были описаны первыми среди клеточных структур в ранних исследованиях микроскопистов. Диаметр ядер обычно равен приблизительно 10 мкм.
Ядро необходимо для жизни клетки, поскольку именно оно регулирует всю ее активность. Связано это с тем, что ядро несет в себе генетическую (наследственную) информацию, заключенную в ДНК. ДНК обладает способностью к репликации, причем ее репликация предшествует делению ядра, так что дочерние ядра также получают ДНК. Деление ядра в свою очередь предшествует клеточному делению, благодаря чему и у всех дочерних клеток имеются ядра. Ядро окружено ядерной оболочкой и содержит хроматин, а также одно или несколько ядрышек.
Ядерная оболочка и ядерные поры
В световом микроскопе мембрана, окружающая ядро, представляется одинарной, однако в действительности это ядерная оболочка, состоящая из двух мембран. Наружная мембрана переходит непосредственно в эндоплазматический ретикулум (ЭР), как это показано на рис. 5.10, 5.11 и 5.24, и, подобно ЭР, может быть усеяна рибосомами, в которых идет синтез белка.

Рис. 5.24. Электронная микрофотография клеточного ядра, полученная с помощью просвечивающего электронного микроскопа. Обратите внимание, что эндоплазматический ретикулум составляет единое целое с ядерной оболочкой.
Ядерная оболочка пронизана ядерными порами (рис. 5.24). Они особенно заметны на препаратах, полученных методом замораживания–травления (рис. 5.25). Через ядерные поры происходит обмен различными материалами между ядром и цитоплазмой, например выход в цитоплазму матричной РНК (мРНК) и рибосомных субчастиц или поступление в ядро рибосомных белков, нуклеотидов и молекул, регулирующих активность ДНК (например, некоторых гормонов). Поры имеют определенную структуру, представляющую собой результат слияния наружной и внутренней мембран ядерной оболочки. Эта структура регулирует прохождение молекул через пору.

Рис. 5.25. Электронная микрофотография ядра, на которой видны ядерные поры. (Препарат получен методом замораживания–травления.) х30 000. При приготовлении таких препаратов быстро замороженные клетки раскалывают металлическим лезвием. Скол проходит в плоскости, представляющей наименьшее сопротивление, часто через мембрану. После удаления льда вытравленная поверхность обнажается.
Хроматин
Хроматин состоит из многих витков ДНК, присоединенных к гистонам — белкам основной природы. Нити ДНК настолько длинны (в каждом ядре соматической клетки человека средняя их длина составляет около 1 м!), что они должны быть как-то упорядоченно упакованы, иначе они перепутаются подобно не смотанной в клубок бечевке. Гистоны и ДНК объединены в структуры, по виду напоминающие бусины; их называют нуклеосомами. Упаковка нуклеосом в хроматине тоже носит регулярный характер.
Слово «хроматин» в переводе означает «окрашенный материал», и назван был так хроматин потому, что он легко окрашивается при подготовке к исследованию с помощью светового микроскопа. Во время деления ядра хроматин окрашивается интенсивнее, а значит, становится и более заметным, что объясняется его конденсацией — образованием более туго скрученных (спирализованных) нитей, которые называются хромосомами. В интерфазе (период между двумя делениями ядра) хроматин переходит в более диспергированное состояние. Часть его, однако, остается плотно спирализованной и по-прежнему интенсивно окрашивается. Эту часть называют гетерохроматином; гетерохроматин имеет вид характерных темных пятен, располагающихся обычно ближе к оболочке ядра (рис. 5.10–13 и рис. 5.24). Остальной, более рыхло спирализованный, хроматин называется эухроматином. Предполагается, что в нем сосредоточена та ДНК, которая в интерфазе генетически активна.
Ядрышко
Ядрышко представляет собой хорошо заметную округлую структуру, находящуюся внутри ядра (рис. 5.12 и 5.24); это — место образования рибосом. В ядре может быть одно или несколько ядрышек. Ядрышко интенсивно окрашивается, потому что оно содержит большое количество ДНК и РНК. РНК близка по своей структуре к ДНК, так как с ДНК она «переписывается» (транскрибируется). В ядрышке имеется особая плотная область, где располагается ДНК одной или нескольких хромосом. Здесь сосредоточено много копий генов, кодирующих рибосомную РНК (рРНК). Во время деления ядра ядрышко становится невидимым, потому что ДНК диспергируется. По завершении деления ядрышко появляется вновь.
Центральную область ядрышка окружает менее плотная периферическая область, где начинается свертывание рибосомной РНК и идет сборка рибосом — РНК соединяется с белком. Не полностью собранные рибосомы переходят по ядерным порам из ядра в цитоплазму, где их сборка завершается.
5.10.2. Цитоплазма
Живое содержимое эукариотических клеток слагается из ядра и цитоплазмы, которые вместе образуют «протоплазму». Цитоплазма состоит из водянистого основного вещества и находящихся в нем разнообразных органелл. Кроме того, в ней присутствуют различные включения — нерастворимые конечные продукты метаболических процессов и запасные вещества.
Растворимая часть цитоплазмы, или ее основное вещество заполняет пространство между клеточными органеллами. Оно содержит систему очень тонких белковых нитей (разд. 5.10.7), в остальном же при изучении в электронном микроскопе представляется прозрачным и бесструктурным. На долю воды в нем приходится приблизительно 90%. В этой воде в растворенном виде содержатся все основные биомолекулы. Истинный раствор образуют ионы и малые молекулы, а именно соли, сахара, аминокислоты, жирные кислоты, нуклеотиды, витамины и растворенные газы. Крупные молекулы — белки — образуют коллоидные растворы. Коллоидным называется раствор, в котором молекулы растворенного вещества относительно велики (см. приложение 1, т. 3). Коллоидный раствор может быть золем (невязким) или гелем (вязким); внешние слои цитоплазмы по своей консистенции часто ближе к гелям.
Основное вещество цитоплазмы — это не только место хранения биомолекул. Здесь же протекают и некоторые метаболические процессы, среди них такой важный процесс, как гликолиз. Если нам удается наблюдать живую цитоплазму, то обычно бросается в глаза ее активность — усиленное движение органелл, в частности митохондрий.
5.10.3. Эндоплазматический ретикулум (ЭР)
Одним из самых важных открытий, сделанных с помощью электронного микроскопа, было обнаружение сложной системы мембран, пронизывающей цитоплазму всех эукариотических клеток. Эта сеть мембран, получившая название эндоплазматический ретикулум (ЭР; от лат. reticulum — сеть), очень хорошо развита в клетке, но лежит за пределами разрешающей способности светового микроскопа. Нередко мембраны усеяны мелкими частицами, которые были названы рибосомами.
На ультратонких срезах ЭР имеет вид множества парных параллельных линий (мембран), располагающихся в цитоплазме (рис. 5.24). Однако иногда срез проходит так, что мы получаем возможность посмотреть как бы сквозь поверхность этих мембран, и тогда можно видеть, что в трех измерениях ЭР имеет не трубчатое, а пластинчатое строение. Модель трехмерной структуры ЭР изображена на рис. 5.26. Видно, что ЭР состоит из уплощенных мембранных мешочков, называемых цистернами. Цистерны ЭР могут быть покрыты рибосомами, и тогда он называется шероховатым или гранулярным ЭР; если рибосомы отсутствуют, то его называют гладким или агранулярным ЭР (строение ближе к трубчатому).
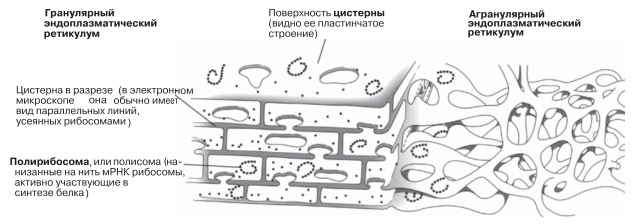
Рис. 5.26. Трехмерная модель эндоплазматического ретикулума.
Функции шероховатого ЭР связаны с транспортом белков, синтезируемых рибосомами на его поверхности. Подробно белковый синтез описан в гл. 23. Здесь достаточно сказать, что растущая белковая молекула, т. е. цепь из аминокислот, или так называемая полипептидная цепь, остается присоединенной к рибосоме до тех пор, пока ее синтез не завершится. Рецептор в мембране ЭР образует канал, по которому новосинтезированный белок переходит в цистерны ЭР.
Транспортируясь по цистернам, белок обычно претерпевает на своем пути весьма существенные изменения. Он может, например, превращаться в гликопротеин. Обычный путь для белка — это путь через шероховатый ЭР в аппарат Гольджи, откуда он либо выходит из клетки наружу (секретируется), либо поступает в другие органеллы той же клетки. Ферменты, содержащиеся в лизосомах, попадают в них именно таким путем (см. также рис. 5.29).
Одной из главных функций агранулярного ЭР является синтез липидов. Так, в эпителии кишечника агранулярный ЭР синтезирует липиды из жирных кислот и глицерола, всасывающихся в кишечнике, а затем передает их в аппарат Гольджи для экспорта. В агранулярном ЭР синтезируются также стероиды — один из классов липидов.
К стероидам принадлежат некоторые гормоны, например кортикостероиды, синтезируемые в коре надпочечников, или половые гормоны тестостерон и эстроген. В мышечных клетках присутствует особая специализированная форма агранулярного ЭР — саркоплазматический ретикулум.
5.10.4. Рибосомы
Рибосомы — это очень мелкие органеллы (диаметром около 20 нм). Число рибосом в цитоплазме живых клеток весьма велико как у прокариот, так и у эукариот. В обычной бактериальной клетке содержится до 10 000 рибосом, а в эукариотических клетках число их во много раз больше.
Рибосомы служат местом синтеза белка. Каждая рибосома состоит из двух субчастиц, как это можно видеть на рис. 5.27. Из-за мелких размеров рибосомы при дифференциальном центрифугировании седиментируют последними среди всех других органелл: рибосомную фракцию можно получить лишь после центрифугирования при 100 000 g в течение 1–2 ч (см. рис. 5.9). Опыты по седиментации выявили существование двух главных типов рибосом, которые были названы 70S- и 80S-рибосомами (S (сведберг) — единица, характеризующая скорость седиментации в центрифуге. Чем больше число S, тем выше скорость седиментации). 70S-рибосомы обнаруживаются у прокариот, а несколько более крупные 80S-рибосомы — в цитоплазме эукариотических клеток. Интересно отметить, что в хлоропластах и митохондриях содержатся 70S-рибосомы, что указывает на какое-то родство этих эукариотических органелл с прокариотами (разд. 7.4.1.).
Рибосомы состоят из примерно равных (по массе) количеств РНК и белка. Входящая в их состав РНК, называемая рибосомной РНК (рРНК), синтезируется в ядрышке. Распределение в рибосоме белковых молекул и молекул РНК показано на рис. 5.27. Вместе те и другие молекулы образуют сложную трехмерную структуру.
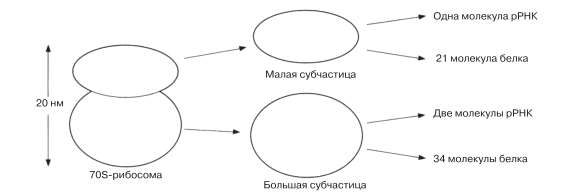
Рис. 5.27. Строение 70S-рибосомы. (В субчастицах 80S-рибосомы больше белка, а в ее большой субчастице содержится не две, а три молекулы рРНК.)
Во время синтеза белка на рибосомах аминокислоты последовательно соединяются друг с другом, формируя полипептидную цепь. Подробно этот процесс описан в гл. 23. Рибосома служит местом связывания для молекул, участвующих в синтезе, т. е. таким местом, где эти молекулы могут занять по отношению друг к другу совершенно определенное положение. В синтезе участвуют: матричная РНК (мРНК), несущая генетические инструкции от клеточного ядра, транспортная РНК (тРНК), доставляющая к рибосоме требуемые аминокислоты, и растущая полипептидная цепь. Кроме того, в этом процессе участвуют факторы инициации, элонгации и терминации цепи. Весь процесс в целом настолько сложен, что без рибосомы он не мог бы идти эффективно (или не шел бы вообще).
В эукариотических клетках отчетливо видны две популяции рибосом — свободные рибосомы и рибосомы, присоединенные к ЭР (рис. 5.24 и 5.26). Строение тех и других идентично, но часть рибосом связана с ЭР через белки, которые они синтезируют. Такие белки обычно секретируются. Примером белка, синтезируемого свободными рибосомами, может служить гемоглобин, образующийся в молодых эритроцитах.
В процессе синтеза белка рибосома перемещается вдоль нитевидной молекулы мРНК. Процесс идет более эффективно, когда вдоль мРНК перемещается не одна рибосома, а одновременно много рибосом, напоминающих в этом случае бусины на нитке. Такие цепи рибосом называются полирибосомами или полисомами. На ЭР полисомы обнаруживаются в виде характерных завитков (рис. 5.26).
5.10.5. Аппарат Гольджи
Структуру, известную теперь как аппарат Гольджи, впервые обнаружил в клетках в 1898 г. Камилло Гольджи (Camillo Golgi), применивший в своих наблюдениях особую методику окрашивания. Однако подробно исследовать ее удалось только с помощью электронного микроскопа. Аппарат Гольджи содержится почти во всех эукариотических клетках и представляет собой стопку уплощенных мембранных мешочков, так называемых цистерн, и связанную с ними систему пузырьков, называемых пузырьками Гольджи.
Трехмерную структуру аппарата Гольджи трудно выявить при изучении ультратонких срезов, однако предполагают, что вокруг центральной стопки формируется сложная система взаимосвязанных трубочек (рис. 5.28).
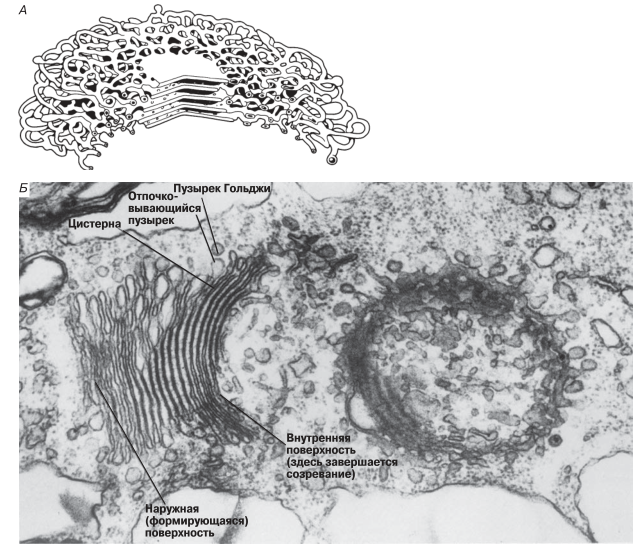
Рис. 5.28. А. Трехмерная структура аппарата Гольджи. Б. Микрофотография, полученная с помощью трансмиссионного электронного микроскопа, на которой видны два аппарата Гольджи: слева — диктиосома в вертикальном разрезе, справа — самая верхняя цистерна, какой она видна сверху (х50 000).
На одном конце стопки постоянно образуются новые цистерны путем слияния пузырьков, отпочковывающихся от агранулярного ЭР. Эта «наружная», или формирующаяся сторона стопки выпуклая, тогда как другая, «внутренняя», где завершается созревание и где цистерны вновь распадаются на пузырьки, имеет вогнутую форму. Стопка состоит из многих цистерн, которые постепенно перемещаются от наружной стороны к внутренней.
Функцию аппарата Гольджи составляют транспорт веществ и химическая модификация поступающих в него клеточных продуктов. Функция эта особенно важна в секреторных клетках, хорошим примером которых могут служить ацинарные клетки поджелудочной железы. Эти клетки секретируют пищеварительные ферменты панкреатического сока в выводной проток железы, по которому они поступают в двенадцатиперстную кишку. На рис. 5.29, А представлена электронная микрофотография такой клетки, а на рис. 5.29, Б — схема упомянутого секреторного пути.
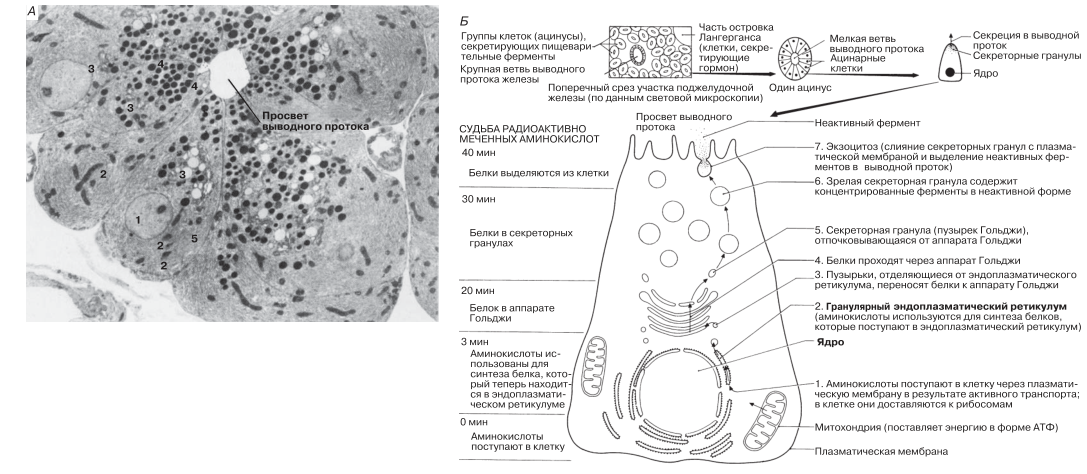
Рис. 5.29. А. Электронная микрофотография ацинуса — группы ацинарных клеток поджелудочной железы, секретирующих ферменты. х10 400. 1 — ядро; 2 — митохондрия; 3 — аппарат Гольджи; 4 — секреторные гранулы; 5 — гранулярный эндоплазматический ретикулум. Б. Схема синтеза и секреции белка (одного из ферментов) в ацинарной клетке поджелудочной железы.
Отдельные этапы этого пути выявляют при помощи радиоактивно меченных аминокислот. Из аминокислот в клетке строятся белки. Используя меченые аминокислоты, можно проследить их включение в белки и передвижение по различным клеточным органеллам. Для этого образцы ткани гомогенизируют через разные промежутки времени после введения аминокислот, разделяют клеточные органеллы центрифугированием (рис. 5.9) и определяют, в каких органеллах наблюдается наивысшая радиоактивность. После концентрирования в аппарате Гольджи белок в пузырьках Гольджи транспортируется к плазматической мембране.
Конечным этапом является секреция неактивного фермента посредством процесса, обратного пиноцитозу. Пищеварительные ферменты, выделяемые поджелудочной железой, синтезируются в неактивной форме, чтобы они не могли разрушать клетки, в которых они образуются.
Примером может служить трипсиноген, превращающийся в активный трипсин в двенадцатиперстной кишке. Обычно у белков, поступающих в аппарат Гольджи из ЭР, имеются короткие олигосахаридные цепи, т. е. они представляют собой гликопротеины (подобно мембранным белкам, изображенным на рис. 5.16). Такие углеводные «антенны» могут претерпевать в аппарате Гольджи модификацию, превращающую их в маркеры, с помощью которых белок направляется строго по своему назначению. Однако, каким образом аппарат Гольджи сортирует и распределяет молекулы, в точности не известно.
Иногда аппарат Гольджи участвует в секреции углеводов, например при синтезе материала клеточных стенок у растений. Рис. 5.30 свидетельствует об усиленной его активности в области «клеточной пластинки», т. е. в той области, где после деления ядра между двумя только что образовавшимися дочерними ядрами закладывается новая клеточная стенка.
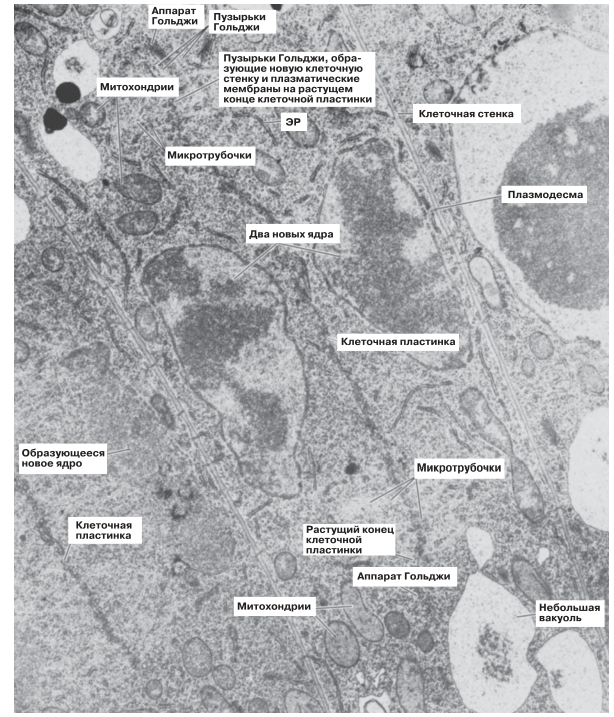
Рис. 5.30. Электронная микрофотография делящейся растительной клетки (располагается на микрофотографии по диагонали). Каждая из двух новых клеток формирует клеточную стенку, материал для которой поступает от центра клетки к периферии. Растущую клеточную стенку называют клеточной пластинкой, поскольку в трехмерном изображении она имеет вид пластинки. Обратите внимание на тесную связь аппарата Гольджи и микротрубочек с растущим концом клеточной пластинки. х15 000.
Пузырьки Гольджи направляются к нужному месту на клеточной пластинке при помощи микротрубочек (разд. 5.10.7). Мембраны этих пузырьков становятся частью плазматических мембран дочерних клеток, а их содержимое используется для построения срединной пластинки и новых клеточных стенок. Целлюлоза поставляется отдельно, но не через аппарат Гольджи, а с помощью микротрубочек. Два рассмотренных нами примера — секреция ферментов клетками поджелудочной железы и образование новых клеточных стенок в делящихся растительных клетках — показывают, каким образом многие клеточные органеллы могут объединяться для выполнения какой-либо одной функции.
Аппаратом Гольджи секретируется гликопротеин муцин, в растворе образующий слизь. Он выделяется бокаловидными клетками, находящимися в толще эпителия слизистой оболочки кишечника и дыхательных путей. В железах листьев некоторых насекомоядных растений, например росянки, аппарат Гольджи секретирует клейкую слизь и ферменты, с помощью которых эти растения ловят и переваривают добычу.
Во многих клетках аппарат Гольджи участвует в секреции слизи, воска, камеди и растительного клея. Помимо секреции различных веществ аппарат Гольджи выполняет и еще одну важную функцию — в нем формируются лизосомы, к описанию которых мы теперь перейдем.
5.10.6. Лизосомы
Лизосомы обнаруживаются почти во всех эукариотических клетках. Они представляют собой простые мембранные мешочки (стенка мешочка состоит из одинарной мембраны), наполненные пищеварительными ферментами — протеазами, нуклеазами и липазами, расщепляющими соответственно белки, нуклеиновые кислоты и липиды. Эти ферменты, катализирующие реакции гидролиза (идущие с присоединением воды), лучше всего работают в кислой среде; содержимое лизосом имеет, следовательно, кислую реакцию. Перечисленные выше ферменты должны быть отделены от всех остальных клеточных компонентов и структур, иначе они разрушат эти компоненты и структуры. В животных клетках лизосомы имеют обычно округлую форму, а диаметр их составляет 0,2–0,5 мкм (рис. 5.31).
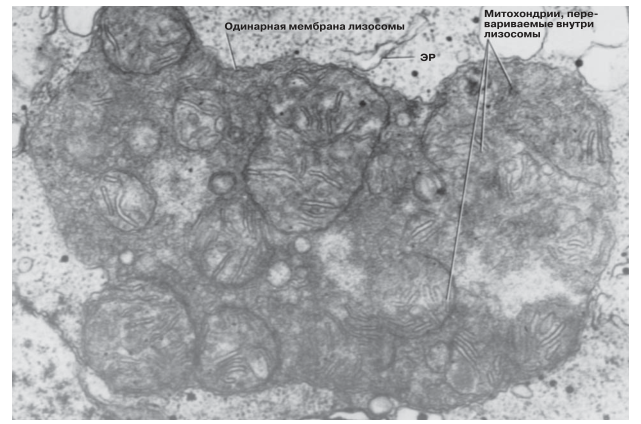
Рис. 5.31.Электронная микрофотография лизосомы, внутри которой перевариваются захваченные ею старые митохондрии. х90 750.
В растительных клетках роль лизосом могут играть крупные центральные вакуоли. Впрочем, в цитоплазме растительных клеток иногда видны тельца, напоминающие по своему виду лизосомы животных клеток.
Заключенные в лизосомах ферменты синтезируются на гранулярном ЭР и транспортируются к аппарату Гольджи. Позже от него отпочковываются пузырьки Гольджи, содержащие ферменты, подвергшиеся необходимым модификациям. Эти пузырьки и превращаются в лизосомы. Лизосомы выполняют ряд функций, которые описаны ниже и перечислены на рис. 5.32.
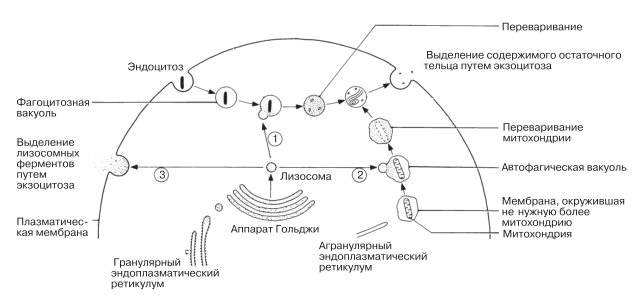
Рис. 5.32. Три процесса, в которых участвуют лизосомы. Цифры 1, 2 и 3 указывают порядок, в котором эти процессы описаны в тексте.
Переваривание материалов, поглощенных посредством эндоцитоза (путь 1, рис. 5.32)
Процесс эндоцитоза описан в разд. 5.9.8. Лизосомы могут сливаться с пузырьками или вакуолями, образующимися в результате эндоцитоза, высвобождать в них свои ферменты и переваривать находящиеся внутри пузырьков или вакуолей материалы. Иногда этот процесс представляет собой способ поглощения пищи, как, например, у некоторых простейших. В других случаях он выполняет защитную функцию, когда, например, специализированные лейкоциты (фагоциты) захватывают и переваривают попавшие в организм бактерии (рис. 5.23).
Продукты переваривания поглощаются и усваиваются цитоплазмой клетки, но часть материала так и остается непереваренной. Вакуоль вместе с этим остаточным материалом направляется к плазматической мембране, и здесь ее содержимое выводится наружу путем экзоцитоза. Своеобразную роль играют лизосомы в клетках щитовидной железы, которые путем пиноцитоза поглощают тиреоглобулин (разд. 5.9.8).
Образовавшиеся пиноцитозные пузырьки сливаются с лизосомами, и тиреоглобулин под действием лизосомных ферментов превращается в активный гормон тироксин. После этого лизосомы путем слияния с плазматической мембраной изливают свое содержимое наружу — выделяют этот гормон в кровь.
Автофагия (путь 2, рис. 5.32)
Автофагией называется процесс, посредством которого клетка поглощает и переваривает внутри лизосом свои собственные не нужные ей структуры. Сначала эти структуры окружаются одинарной мембраной, отделяющейся обычно от агранулярного эндоплазматического ретикулума, а затем такой мембранный мешочек с заключенной в нем структурой сливается с лизосомой, образуя так называемую автофагическую вакуоль, в которой структура переваривается. Данная последовательность событий входит как составная часть в естественный круговорот цитоплазматических органелл, при котором старые органеллы заменяются новыми.
Выделение ферментов из клетки (экзоцитоз) (путь 3, рис. 5.32)
Иногда ферменты, содержащиеся в лизосомах, высвобождаются из клетки наружу. Это происходит, например, в процессе развития организма при замене хрящей костной тканью. Аналогичное явление можно наблюдать, когда основное вещество кости разрушается при перестройке костной ткани в ответ на повреждения, при новой нагрузке и т. п. В спермиях содержатся специализированные лизосомы, называемые акросомами. Из них наружу непосредственно перед оплодотворением выделяются ферменты, с помощью которых спермий прокладывает себе путь к яйцеклетке сквозь окружающие ее клеточные слои, переваривая отделяющий его от яйцеклетки материал.
Автолиз
Автолиз — это саморазрушение клетки, наступающее в результате высвобождения содержимого ее лизосом внутри самой клетки. Именно в связи с этим лизосомы были в свое время метко названы «орудиями самоубийства» (suicide bags). При некоторых процессах дифференцировки автолиз представляет собой нормальное явление; он может распространяться на всю ткань, как это, например, имеет место при резорбции хвоста головастика во время метаморфоза.
Другой пример — превращения, которые претерпевает матка. Во время беременности матка сильно увеличивается, чтобы вместить растущий плод. После родов она постепенно приобретает свои обычные размеры в результате самопереваривания большого числа клеток. Наблюдается автолиз также и в мышцах долго остающихся без работы. После гибели клетки тоже наступает автолиз; именно поэтому пищевые продукты портятся, если они не были заморожены. Иногда автолиз является следствием некоторых лизосомных болезней.
5.10.7. Микротрубочки
Электронный микроскоп выявил наличие структуры в «основном веществе» цитоплазмы, которое ранее представлялось бесструктурным. Во всех эукариотических клетках была обнаружена сеть тонких белковых нитей. Все вместе они образуют так называемый цитоскелет. Различают по меньшей мере три типа таких структур: микротрубочки, микрофиламенты и промежуточные филаменты. Их функции связаны с внутриклеточным движением, со способностью клеток поддерживать свою форму, а также с некоторыми другими видами активности клеток, такими, например, как эндоцитоз и экзоцитоз. Мы рассмотрим здесь только микротрубочки.
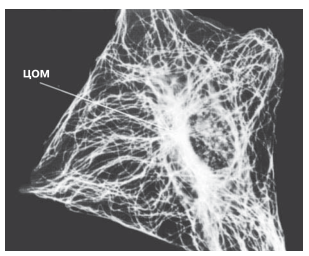
Рис. 5.33. Распределение микротрубочек в клетке. Микротрубочки расходятся от центра организации микротрубочек (ЦОМ), находящегося рядом с ядром. В ЦОМ содержится центриоль. Микротрубочки видны на этой микрофотографии благодаря использованию флуоресцирующих антител, способных специфически соединяться с их белком. Представленная здесь клетка — фибробласт; фибробласты обычно содержатся в соединительной ткани; в них синтезируется коллаген.
Микротрубочки содержатся почти во всех эукариотических клетках (рис. 5.33). Это полые, очень тонкие неразветвленные трубочки диаметром приблизительно 24 нм; их стенки толщиной около 5 нм построены из спирально упакованных субъединиц белка тубулина (рис. 5.34).
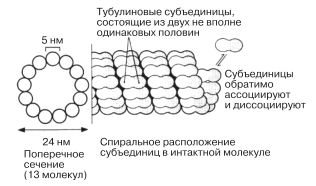
Рис. 5.34. Расположение тубулиновых субъединиц в микротрубочке.
Рис. 5.30 дает представление о том, как выглядят микротрубочки на электронных микрофотографиях. Растут микротрубочки с одного конца путем добавления тубулиновых субъединиц. Рост видимо, может начаться лишь при наличии матрицы; есть основания полагать, что роль таких матриц играют какие-то очень мелкие кольцевые структуры, которые были выделены из клеток и которые, как выяснилось, состоят из тубулиновых субъединиц. В интактных клетках ту же функцию выполняют центриоли, поэтому их иногда называют центрами организации микротрубочек (ЦОМ). Центриоли состоят из коротких микротрубочек. Микротрубочки принимают участие в различных внутриклеточных процессах; некоторые мы здесь упомянем.
Центриоли и деление ядра
Центриоли это мелкие полые цилиндры (длиной 0,3–0,5 мкм и около 0,2 мкм в диаметре), встречающиеся в виде парных структур почти во всех животных клетках. Каждая центриоль построена из девяти триплетов микротрубочек. В начале деления ядра центриоли удваиваются и две новые пары центриолей расходятся к полюсам веретена — структуры, по экватору которой выстраиваются перед своим расхождением хромосомы. Само веретено состоит из микротрубочек («нитей веретена»), при сборке которых центриоли играют роль центров организации. Микротрубочки регулируют расхождение хроматид или хромосом. Осуществляется это за счет скольжения микротрубочек (гл. 23). В клетках высших растений центриоли отсутствуют, хотя веретено в них при делении ядра образуется.
Возможно, что в этих клетках имеются какие-то очень мелкие центры организации микротрубочек, не выявляемые даже при помощи электронного микроскопа.
Базальные тельца, реснички и жгутики
Реснички и жгутики идентичны по своему строению, но жгутики длиннее ресничек. Обе эти органеллы представляют собой выросты клеток. Движутся они либо однонаправленно (биение ресничек), либо волнообразно (движения жгутиков). Служат реснички и жгутики как для передвижения отдельных клеток, так и для того, чтобы перегонять жидкость вдоль поверхности клеток (так перегоняют реснички слизь в дыхательных путях). В основании каждой реснички и жгутика всегда обнаруживается базальное тельце.
По своему строению базальные тельца идентичны центриолям и можно думать, что они образуются путем удвоения центриолей. Вероятно, они также действуют как центры организации микротрубочек, потому что ресничкам и жгутикам тоже свойственно характерное расположение микротрубочек («9 + 2»; гл. 18).
В ресничках и жгутиках движение осуществляется за счет скольжения микротрубочек. Более подробно эти процессы описаны в гл. 18. Отметим, что жгутики бактерий устроены проще, чем жгутики эукариот, и базальные тельца у них отсутствуют.
Внутриклеточный транспорт
Микротрубочки участвуют также в перемещении различных клеточных органелл, например в перемещении пузырьков Гольджи к формирующейся клеточной пластинке (рис. 5.30). В клетке идет непрерывный транспорт: перемещаются пузырьки Гольджи, направляются к аппарату Гольджи пузырьки, отпочковывающиеся от ЭР, движутся лизосомы, митохондрии и другие органеллы. Все это движение приостанавливается, если повреждена система микротрубочек.
Цитоскелет
Микротрубочки выполняют в клетках еще и структурную роль: эти длинные, трубчатые, достаточно жесткие структуры образуют опорную систему клетки, являясь частью цитоскелета. Они способствуют определению формы клеток в процессе дифференцировки и поддержанию формы дифференцированных клеток; нередко они располагаются в зоне, непосредственно примыкающей к плазматической мембране (рис. 5.10 и 5.11). Животные клетки, в которых система микротрубочек повреждена, принимают сферическую форму. В растительных клетках расположение микротрубочек точно соответствует расположению целлюлозных волокон, отлагающихся при построении клеточной стенки; таким образом микротрубочки косвенно определяют форму клетки.
5.10.8. Микроворсинки
Микроворсинками называют пальцевидные выросты плазматической мембраны некоторых животных клеток (рис. 5.10 и 5.12). Иногда микроворсинки увеличивают площадь поверхности клетки в 25 раз, поэтому они особенно многочисленны на поверхности клеток всасывающего типа, а именно в эпителии тонкого кишечника и извитых канальцев нефронов. Это увеличение площади всасывающей поверхности способствует и лучшему перевариванию пищи в кишечнике, потому что некоторые пищеварительные ферменты находятся на поверхности клеток и связаны с ней (разд. 8.3.8).
Бахрома микроворсинок на эпителиальных клетках хорошо видна в световом микроскопе; это так называемая щеточная каемка эпителия. В каждой микроворсинке содержатся пучки актиновых и миозиновых нитей. Актин и миозин — это белки мышц, участвующие в мышечном сокращении. В основании микроворсинок актиновые и миозиновые нити, связываясь с нитями соседних микроворсинок, образуют сложную сеть. Вся эта система в целом поддерживает микроворсинки в расправленном состоянии и позволяет им сохранять свою форму, обеспечивая в то же время и скольжение актиновых нитей вдоль миозиновых (наподобие того, как это происходит при мышечном сокращении).
5.10.9. Митохондрии
Митохондрии содержатся во всех аэробных эукариотических клетках. Об их структуре и функции дает некоторое представление рис. 5.12. Главную функцию митохондрий составляет аэробное дыхание, поэтому они подробно описаны в разд. 9.3.
5.10.10. Клеточные стенки
Растительные клетки, подобно клеткам прокариот и грибов, заключены в сравнительно жесткую клеточную стенку, материал для построения которой секретирует сама находящаяся в ней живая клетка (протопласт). По своему химическому составу клеточные стенки растений отличаются от клеточных стенок прокариот и грибов (табл. 2.2).
Клеточная стенка, отлагающаяся во время деления клеток растения, называется первичной клеточной стенкой. Позже в результате утолщения она может превратиться во вторичную клеточную стенку. На рис. 5.30 воспроизведена электронная микрофотография, на которой можно видеть одну из ранних стадий этого процесса.
Строение клеточной стенки
Первичная клеточная стенка состоит из целлюлозных фибрилл, погруженных в матрикс, в состав которого входят другие полисахариды. Целлюлоза тоже представляет собой полисахарид (ее химическое строение описано в разд. 3.2.3.). Она обладает высокой прочностью на разрыв, сравнимой с прочностью стали. Матрикс состоит из полисахаридов, которые для удобства описания делят обычно на пектины и гемицеллюлозы. Пектины — это кислые полисахариды с относительно высокой растворимостью. Срединная пластинка, скрепляющая стенки соседних клеток, состоит из клейких студнеобразных пектатов (солей пектина) магния и кальция.
Гемицеллюлозы — это смешанная группа полисахаридов, растворимых в щелочах. У гемицеллюлоз, как и у целлюлозы, молекулы имеют форму цепи, однако их цепи короче, менее упорядочены и сильнее разветвлены. Клеточные стенки гидратированы: 60–70% их массы обычно составляет вода. По свободному пространству клеточной стенки вода перемещается беспрепятственно.
У некоторых клеток, например у клеток мезофилла листа, на всем протяжении их жизни имеется только первичная клеточная стенка. Однако у большинства клеток на внутреннюю поверхность первичной клеточной стенки (снаружи от плазматической мембраны) отлагаются дополнительные слои целлюлозы, т. е. возникает вторичная клеточная стенка. В любом слое вторичного утолщения целлюлозные волокна располагаются под одним и тем же углом, но в разных слоях этот угол различен, чем и обеспечивается еще большая прочность структуры. Такое расположение целлюлозных волокон показано на рис. 5.35.

Рис. 5.35. Электронная микрофотография, на которой видны целлюлозные волокна в отдельных слоях клеточной стенки зеленой морской водоросли Chaetomorpha melagonium. Толщина целлюлозных микрофибрилл составляет 20 нм. Для получения контрастного изображения произведено напыление сплавом платины с золотом.
Некоторые клетки, такие, как трахеальные элементы ксилемы и клетки склеренхимы, претерпевают интенсивную лигнификацию (одревеснение). При этом все слои целлюлозы пропитываются лигнином — сложным полимерным веществом, не относящимся к полисахаридам. Клетки протоксилемы лигнифицируются лишь частично. В других случаях лигнификация бывает сплошной, если не считать так называемых поровых полей, т. е. тех участков в первичной клеточной стенке, через которые осуществляется контакт между соседними клетками при помощи группы плазмодесм (разд. 6.1.3 и рис. 6.8).
Лигнин скрепляет целлюлозные волокна и удерживает их на месте. Он действует как очень твердый и жесткий матрикс, усиливающий прочность клеточных стенок на растяжение и в особенности на сжатие (предотвращает прогибы). Это главный опорный материал дерева. Он также предохраняет клетки от повреждения под действием физических и химических факторов. Вместе с целлюлозой, остающейся в клеточных стенках, лигнин придает древесине те особые свойства, которые делают ее незаменимым строительным материалом.
| Дополнение. Композиционные материалы
Материалы с высокой механической прочностью состоящие, подобно клеточным стенкам более чем из одного компонента, называются композиционными материалами или композитами. Прочность таких материалов обычно выше, чем у каждого из компонентов в отдельности. Системы из матрицы, армированной волокнами, используются весьма широко, поэтому изучение их свойств составляет важный раздел как в новейшей технике, так и в биологии. Матрица композита передает приложенную к ней нагрузку волокнам, обладающим высокой прочностью на растяжение. Она же повышает сопротивление сжатию и срезывающему усилию. Примером композита может служить такой известный строительный материал, как железобетон, в котором роль матрицы играет бетон, а роль упрочнителя — стальная арматура. Из более современных и более легких структурных материалов можно назвать, например, стеклопластики и углепластики, в которых пластиковая матрица армирована соответственно стеклянными или углеродными волокнами. К жестким композиционным материалам биологического происхождения относятся древесина, кость, хрящ и материал экзоскелета членистоногих, а к гибким — некоторые типы соединительной ткани и кожа. |
Функции клеточной стенки
Ниже перечислены основные функции клеточных стенок растений.
1. Клеточные стенки обеспечивают отдельным клеткам и растению в целом механическую прочность и опору. В некоторых тканях прочность усиливается благодаря интенсивной лигнификации (небольшое количество лигнина присутствует во всех клеточных стенках). Особо важную роль играет лигнификация клеточных стенок у древесных и кустарниковых пород.
2. Относительная жесткость клеточных стенок и сопротивление растяжению обусловливают тургесцентность клеток, когда в них осмотическим путем поступает вода. Это усиливает опорную функцию во всех растениях и служит единственным источником опоры для травянистых растений и для таких органов, как листья, т. е. там, где отсутствует вторичный рост. Клеточные стенки также предохраняют клетки от разрыва в гипотонической среде.
3. Ориентация целлюлозных микрофибрилл ограничивает и в известной мере регулирует как рост, так и форму клеток, поскольку от расположения этих микрофибрилл зависит способность клеток к растяжению. Если, например, микрофибриллы располагаются поперек клетки, опоясывая ее как бы обручами, то клетка, в которую путем осмоса поступает вода, будет растягиваться в продольном направлении.
4. Система связанных друг с другом клеточных стенок (апопласт) служит главным путем, по которому передвигаются вода и растворенные в ней питательные вещества (гл. 13). Клеточные стенки скреплены между собой с помощью срединных пластинок. В стенках имеются небольшие поры, сквозь которые проходят цитоплазматические тяжи, называемые плазмодесмами. Плазмодесмы связывают живое содержимое отдельных клеток, т. е. объединяют все протопласты в единую систему, в так называемый симпласт (гл. 13).
5. Наружные клеточные стенки эпидермальных клеток покрываются особой пленкой — кутикулой, состоящей из воскообразного вещества кутина, что снижает потери воды и уменьшает риск проникновения в растение болезнетворных организмов. В пробковой ткани клеточные стенки по завершении вторичного роста пропитываются суберином, выполняющим сходную функцию.
6. Клеточные стенки сосудов ксилемы и ситовидных трубок флоэмы приспособлены для дальнего транспорта веществ по растению. Этот вопрос рассматривается в гл. 6 и 13.
7. Стенки клеток эндодермы корня пропитаны суберином и поэтому служат барьером на пути движения воды (гл. 13).
8. У некоторых клеток их видоизмененные стенки хранят запасы питательных веществ; таким способом, например, запасаются гемицеллюлозы в некоторых семенах.
9. У передаточных клеток площадь поверхности клеточных стенок увеличена и соответственно увеличена площадь поверхности плазматической мембраны, что повышает эффективность переноса веществ путем активного транспорта (гл. 13).
5.10.11. Плазмодесмы
Плазмодесмы — это живые связи, соединяющие соседние клетки растения через очень мелкие поры в смежных клеточных стенках (рис. 5.13). Плазматические мембраны соседних клеток переходят непосредственно одна в другую, выстилая поры. Через просвет каждой поры переходит из клетки в клетку и агранулярный эндоплазматический ретикулум. Такая система упрощает связи и координацию между отдельными растительными клетками, поскольку ионам и молекулам не приходится преодолевать на своем пути плазматическую мембрану. Их передвижение, однако, регулируется. Вирусы способны использовать поры в клеточных стенках и могут переходить из клетки в клетку по плазмодесмам. Плазмодесмы играют также роль при формировании пор в ситовидных пластинках флоэмы.
5.10.12. Вакуоли
Вакуоль представляет собой наполненный жидкостью мембранный мешок, стенка которого состоит из одинарной мембраны. В животных клетках содержатся относительно небольшие вакуоли: фагоцитозные, пищеварительные, автофагические и сократительные. Иная картина наблюдается в растительных клетках, особенно в зрелой паренхиме. Здесь клетки содержат одну большую центральную вакуоль, окруженную мембраной, которая носит название тонопласта (рис. 5.11). Жидкость, заполняющая центральную вакуоль, называется клеточным соком.
Это концентрированный раствор, содержащий минеральные соли, сахара, органические кислоты, кислород, диоксид углерода, пигменты и некоторые отходы жизнедеятельности или «вторичные» продукты метаболизма. Ниже перечислены функции, выполняемые вакуолями.
1. Вода обычно поступает в концентрированный клеточный сок путем осмоса, через избирательно проницаемый тонопласт. В результате в клетке развивается тургорное давление и цитоплазма прижимается к клеточной стенке. Осмотическое поглощение воды играет важную роль при растяжении клеток во время роста, а также в общем водном режиме растения.
2. Иногда в вакуолях содержатся растворимые пигменты. В эту группу входят антоцианины, имеющие красную, синюю или пурпурную окраску, и некоторые родственные соединения, окрашенные в желтый или кремовый цвет. Именно эти пигменты главным образом и определяют окраску цветков (например, у роз, фиалок и георгин), а также окраску плодов, почек и листьев. У листьев они обусловливают различные оттенки осенней окраски, которая зависит также от фотосинтетических пигментов, содержащихся в хлоропластах. Окраска играет роль в привлечении насекомых, птиц и некоторых других животных, участвующих в опылении растений и в распространении семян.
3. У растений в вакуолях иногда содержатся гидролитические ферменты и тогда вакуоли функционируют как лизосомы. После гибели клетки тонопласт, как и все другие мембраны, теряет свою избирательную проницаемость, и ферменты высвобождаются из вакуолей, вызывая автолиз.
4. В вакуолях растения могут накапливаться конечные и некоторые вторичные продукты метаболизма. Из конечных продуктов иногда обнаруживаются, например, кристаллы оксалата кальция. Вторичные продукты, в частности алкалоиды и таннины, выполняют, возможно, защитную функцию, предотвращая поедание таких растений травоядными животными. Может накапливаться в вакуолях и латекс — млечный сок растений, такой, например, как у одуванчика. В млечном соке бразильской гевеи содержатся соединения, из которых синтезируют каучук, а в млечном соке мака снотворного — такие алкалоиды, как морфин, из которого получают героин.
5. Некоторые растворимые компоненты клеточного сока, например сахароза и минеральные соли, играют роль запасных питательных веществ, при необходимости используемых цитоплазмой.
5.10.13. Хлоропласты
Хлоропласты — это пластиды, содержащие хлорофилл и каротиноиды и осуществляющие фотосинтез. Они находятся главным образом в листьях. Описание их дано в разд. 7.4.1.