5.9. Клеточные мембраны
Биология — Тейлор Д., Грин Н., Стаут У. 1 том — 2013
Клеточные мембраны играют важную роль по ряду причин. Они отделяют клеточное содержимое от внешней среды, регулируют обмен между клеткой и средой (поступление в клетку питательных веществ и удаление из нее «отходов») и делят клетки на отсеки, или компартменты, предназначенные для тех или иных метаболических путей, например для фотосинтеза или аэробного дыхания. Некоторые химические реакции, в частности световые реакции фотосинтеза в хлоропластах, протекают на самих мембранах. Здесь же на мембранах располагаются и рецепторные участки для распознавания гормонов, нейромедиаторов или других химических веществ, поступающих из окружающей среды или из других частей самого организма. Знакомство со всеми свойствами клеточных мембран необходимо для понимания того, как функционирует клетка.
5.9.1. Мембраны обладают избирательной проницаемостью
С конца прошлого века известно, что клеточные мембраны ведут себя не так, как полупроницаемые мембраны, способные пропускать лишь воду и другие малые молекулы, например молекулы газов. Клеточные мембраны обладают избирательной проницаемостью: через них медленно диффундируют глюкоза, аминокислоты, жирные кислоты, глицерол и ионы, причем сами мембраны активно регулируют этот процесс — одни вещества пропускают, а другие нет.
5.9.2. Мембраны содержат белки и липиды
Ранние работы по проницаемости мембран показали, что органические растворители, например спирт, эфир или хлороформ, проникают через мембраны даже быстрее, чем вода. Это свидетельствовало о том, что в мембранах есть какая-то неполярная часть; иными словами, что мембраны содержат липиды. Позже данное предположение удалось подтвердить химическим анализом. Выяснилось, что мембраны состоят почти целиком из белков и липидов. О белках мы будем говорить ниже. Липиды в мембранах представлены преимущественно фосфолипидами.
5.9.3. Фосфолипиды
Подробно структура фосфолипидов описана в гл. 3 (рис. 3.18). Молекула фосфолипида состоит из полярной 1 головы (фосфатной группы) и двух неполярных углеводородных хвостов (остатков жирных кислот). Полярность означает, что в молекуле неравномерно распределены заряды, и это делает ее растворимой в воде. Характерная особенность фосфолипидов состоит в том, что голова у них гидрофильна, а хвосты гидрофобны.
Небольшое количество фосфолипида, растекаясь по поверхности воды, образует монослой, как это видно на рис. 5.14. Неполярные углеводородные хвосты выступают из воды, а полярные гидрофильные головы лежат на ее поверхности.

Рис. 5.14. Монослой фосфолипидных молекул на поверхности воды.
Если фосфолипида больше, чем нужно для того, чтобы просто покрыть поверхность воды, или если взболтать фосфолипид с водой, то образуются частицы, которые называются мицеллами. Гидрофобные хвосты фосфолипидных молекул упрятаны в них внутрь и тем самым защищены от контакта с водой (рис. 5.15,А). На рис. 5.15,Б и 5.15,В изображены структуры, в которых имеется двойной слой фосфолипидных молекул — так называемый бислой. Теперь известно, что фосфолипидный бислой — основная структура клеточных мембран.

Рис. 5.15. Сферическая мицелла (А) и двойной слой (бислой) фосфолипидных молекул (Б), представленные в разрезе. В. Трехмерная модель бислоя.
5.9.4. Белки
С помощью метода замораживания–скалывания удалось выяснить, как встроены в фосфолипидный бислой белки. Этот метод предполагает быстрое замораживание, а затем расщепление мембран с помощью очень острого лезвия. При этом их внутренние поверхности обнажаются и становятся доступными для обозрения. На поверхностях обнаруживаются различные частицы, главным образом белковые, которые иногда погружены в бислой, а иногда пронизывают его насквозь. В целом, чем выше метаболическая активность мембраны, тем больше в ней обнаруживается белковых частиц: в мембранах хлоропластов (75% белка) их очень много (рис. 7.12), а в метаболически инертной оболочке аксона (18% белка) их нет совсем. Неодинаково распределены они также на внутренней и наружной сторонах мембраны.
5.9.5. Гликолипиды и холестерол
В мембранах содержатся также гликолипиды и холестерол. Гликолипиды — это липиды с присоединенными к ним углеводами. Как и у фосфолипидов, у гликолипидов имеются полярные головы и неполярные хвосты. Холестерол близок к липидам; в его молекуле также имеется полярная часть.
5.9.6. Жидкостно-мозаичная модель мембраны
В 1972 г. Сингер и Николсон (Singer, Nicolson) предложили жидкостно-мозаичную модель мембраны, согласно которой белковые молекулы плавают в жидком фосфолипидном бислое. Они образуют в нем как бы своеобразную мозаику, но поскольку бислой этот жидкий, то и сам мозаичный узор не жестко фиксирован; белки могут менять в нем свое положение. Покрывающая клетку тонкая мембрана напоминает пленку мыльного пузыря — она тоже все время «переливается». На рис. 5.16 представлено плоскостное изображение жидкостно-мозаичной модели мембраны и ее трехмерная модель.

Рис. 5.16. А. Трехмерное изображение жидкостно-мозаичной модели мембраны. Б. Плоскостное ее изображение.
Гликопротеины и гликолипиды связаны только с наружной поверхностью мембраны.
Ниже суммированы известные нам данные, касающиеся строения и свойств клеточных мембран.
- Толщина мембран составляет около 7 нм.
- Основная структура мембраны — фосфолипидный бислой.
- Гидрофильные головы фосфолипидных молекул обращены наружу — в сторону водного содержимого клетки и в сторону наружной водной среды.
- Гидрофобные хвосты обращены внутрь — они образуют гидрофобную внутреннюю часть бислоя.
- Фосфолипиды находятся в жидком состоянии и быстро диффундируют внутри бислоя — перемещаются в латеральном направлении.
- Жирные кислоты, образующие хвосты фосфолипидных молекул, бывают насыщенными и ненасыщенными (см. гл. 3, рис. 3.17). В ненасыщенных кислотах имеются изломы, что делает упаковку бислоя более рыхлой. Следовательно, чем больше степень ненасыщенности, тем более жидкую консистенцию имеет мембрана.
- Большая часть белков плавает в жидком фосфолипидном бислое, образуя в нем своеобразную мозаику, постоянно меняющую свой узор.
- Белки сохраняют связь с мембраной, поскольку в них есть участки, состоящие из гидрофобных аминокислот, взаимодействующих с гидрофобными хвостами фосфолипидов; вода из этих мест выталкивается. Другие участки белков гидрофильны. Они обращены либо к окружению клетки, либо к ее содержимому, т. е. к водной среде.
- Некоторые мембранные белки лишь частично погружены в фосфолипидный бислой, тогда как другие пронизывают его насквозь.
- К некоторым белкам и липидам присоединены разветвленные олигосахаридные цепочки, играющие роль антенн. Такие соединения называются соответственно гликопротеинами и гликолипидами.
- В мембранах содержится также холестерол. Подобно ненасыщенным жирным кислотам он нарушает плотную упаковку фосфолипидов и делает их более жидкими. Это важно для организмов, живущих в холодной среде, где мембраны могли бы затвердевать. Холестерол делает мембраны также более гибкими и вместе с тем более прочными. Без него они бы легко разрывались.
- Две стороны мембраны, наружная и внутренняя, различаются и по составу, и по функциям.
5.9.7. Функции мембран
Фосфолипидный бислой, как уже было сказано, составляет основу структуры мембраны. Он также ограничивает проникновение полярных молекул и ионов в клетку и выход их из нее. Ряд функций выполняют и другие компоненты мембран.
- Белки-каналы и белки-переносчики осуществляют избирательный транспорт полярных молекул и ионов через мембрану (об активном транспорте и облегченной диффузии см. разд. 5.9.8).
- Ферменты. Белки нередко функционируют как ферменты. В качестве примера укажем на микроворсинки эпителия, выстилающего некоторые отделы кишечника. Плазматические мембраны этих эпителиальных клеток содержат пищеварительные ферменты.
- Рецепторные молекулы. У всех белковых молекул весьма специфическая конформация, о чем мы уже говорили в гл. 3 и 4. Это делает их идеальными рецепторами, т. е. молекулами, при помощи которых от клетки к клетке передаются те или иные сигналы. Например, гормоны, являющиеся химическими посредниками, циркулируют в крови, но присоединяются они только к особым клеткам-мишеням, у которых есть соответствующие рецепторы. Нейромедиаторы — химические вещества, обеспечивающие проведение нервных импульсов, — тоже связываются с особыми рецепторными белками нервных клеток.
- Антигены действуют как маркеры, своего рода «ярлыки», позволяющие опознать клетку. Это гликопротеины, т. е. белки с присоединенными к ним разветвленными олигосахаридными боковыми цепями, играющими роль «антенн». Существует бесчисленное множество возможных конфигураций этих боковых цепей, так что у каждой клетки может быть свой особый маркер. С помощью маркеров клетки способны распознавать другие клетки и действовать согласованно с ними, например при формировании тканей и органов у многоклеточных организмов. Это же свойство позволяет иммунной системе распознавать и атаковать чужеродные антигены.
- У гликолипидов тоже имеются разветвленные олигосахаридные боковые цепи и они также помогают клеткам распознавать друг друга. Гликолипиды могут служить рецепторами для химических сигналов. Вместе с гликопротеинами гликолипиды обеспечивают правильное сцепление клеток при их объединении в ткани.
- Перенос энергии. При фотосинтезе и дыхании в мембранах соответственно хлоропластов и митохондрий действуют системы переноса энергии, в которых также участвуют белки.
- Холестерол служит дополнительным «стопором», препятствующим перемещению полярных молекул через мембрану в обоих направлениях — в клетку и из клетки.
5.9.8. Транспорт через плазматическую мембрану
Хотя толщина плазматических мембран составляет обычно всего около 7 нм, они служат барьером для ионов и молекул, в особенности для полярных (водорастворимых) молекул, таких как глюкоза или аминокислоты, поскольку неполярные (гидрофобные) липиды мембран эти вещества отталкивают. Барьер не дает водному содержимому клетки ускользнуть из нее.
Тем не менее по ряду причин транспорт через мембраны все же должен идти, поскольку необходимо обеспечивать:
- доставку питательных веществ;
- удаление конечных продуктов обмена («отходов»);
- секрецию различных полезных веществ;
- создание ионных градиентов, весьма важных для нервной и мышечной деятельности;
- поддержание в клетке соответствующего pH и надлежащей ионной концентрации, которые нужны для эффективной работы клеточных ферментов.
Мы обсудим здесь транспорт веществ через плазматическую мембрану, отметив, что аналогичный характер носит и транспорт через мембраны клеточных органелл. Существует четыре основных механизма для поступления веществ в клетку или выхода их из клетки наружу: диффузия, осмос, активный транспорт и экзо- или эндоцитоз. Два первых процесса носят пассивный характер, т. е. не требуют затрат энергии; два последних — активные процессы, связанные с потреблением энергии.
Диффузия и облегченная диффузия
Диффузией называют перемещение веществ из области с высокой их концентрацией в область с низкой концентрацией по диффузионному градиенту. Это пассивный процесс, не требующий затрат энергии и протекающий спонтанно.
Если, например, оставить в закрытой комнате открытым флакон духов, то духи будут постепенно распространяться по всей комнате до тех пор, пока не распределятся в ней равномерно. Обусловливается это беспорядочным движением молекул за счет их кинетической энергии (энергии движения). Каждый тип молекул перемещается по своему собственному диффузионному градиенту независимо от других молекул. Кислород, например, диффундирует из легких в кровь, а диоксид углерода — в обратном направлении.
На скорость диффузии влияют в первую очередь три фактора.
- Крутизна диффузионного градиента, т. е. различие в концентрации между пунктом А и пунктом В; чем круче градиент, тем выше скорость диффузии. Клетке выгодно поддерживать крутой диффузионный градиент, если требуется быстрая доставка тех или иных веществ. В легких, например, этого можно достичь за счет ускорения тока крови, проходящей через них, или за счет усиленного дыхания.
- Чем больше площадь поверхности мембраны, через которую диффундирует вещество, тем быстрее идет диффузия. Для клеток, форма которых близка к сферической, площадь поверхности по отношению к объему тем меньше, чем крупнее клетка. Это налагает ограничения на размеры клеток. Очень крупная аэробная клетка не могла бы, например, достаточно быстро получать кислород, если бы он поступал в нее только за счет диффузии. Некоторые животные клетки для увеличения площади поверхности, через которую идет поглощение, снабжены микроворсинками.
- Скорость диффузии быстро снижается с увеличением расстояния (она обратно пропорциональна квадрату расстояния). Диффузия, следовательно, эффективна лишь на очень коротких отрезках пути. Это тоже налагает ограничения на размеры клеток. Диффузия служит клеткам для внутреннего транспорта молекул, поэтому диаметр большинства клеток не превышает 50 мкм и любая часть клетки отстоит от ее поверхности не более, чем на 25 мкм. Какая-либо аминокислота может, например, преодолеть путь в несколько микрометров за считанные секунды, но чтобы пройти несколько сантиметров, ей понадобится не один день. С этой точки зрения важно, что мембраны такие тонкие — молекулы или ионы могут проходить через них быстро.
Факторы, влияющие на скорость диффузии, объединены в законе Фика. Он гласит, что скорость диффузии пропорциональна следующему выражению:

Итак, какие молекулы могут проходить через мембраны за счет диффузии? Быстро диффундируют через мембраны такие газы, как кислород и диоксид углерода. Молекулы воды, хотя и сильно поляризованные, достаточно малы для того, чтобы без помех проскользнуть между гидрофобными молекулами фосфолипидов. Вместе с тем ионы и более крупные полярные молекулы гидрофобными участками мембраны отталкиваются, а потому диффундируют через мембрану крайне медленно. Для их поступления в клетку требуются другие механизмы.
Некоторые ионы и полярные молекулы проникают в клетку при помощи особых транспортных белков. Это белки-каналы и белки-переносчики. Заполненные водой гидрофильные каналы, или поры, этих белков имеют строго определенную форму, соответствующую тому или иному иону или молекуле. Иногда канал проходит не внутри одной белковой молекулы, а между несколькими соседними молекулами. Диффузия по каналам идет в обоих направлениях. Такую диффузию — при помощи транспортных белков — называют облегченной диффузией. Транспортные белки, по которым проходят ионы, называются ионными каналами. Обычно ионные каналы снабжены «воротами», т. е. могут открываться и закрываться. Ионные каналы, способные открываться и закрываться, играют важную роль при проведении нервных импульсов.
У белков-каналов форма фиксирована (рис. 5.16, Б). Было показано, что болезнь, известная как кистозный фиброз, есть результат дефекта в белке, который служит каналом для хлорид-ионов. У белков-переносчиков форма, наоборот, претерпевает быстрые изменения, до 100 циклов в секунду. Они существуют в двух состояниях, и механизм их действия напоминает игру в «пинг-понг». На рис. 5.17 показано, как функционирует этот механизм. Связывающие участки белка-переносчика в одном состоянии («пинг») обращены наружу, а в другом («понг») внутрь клетки. Чем выше концентрация растворенных молекул или ионов, тем больше шансов на то, что они окажутся связанными. Если концентрация растворенного вещества снаружи выше, чем в клетке, как в примере с глюкозой на рис. 5.17, то реальный поток этого вещества будет направлен внутрь, и оно будет поступать в клетку. Именно так глюкоза проникает в эритроциты. Перемещение такого рода имеет все характерные признаки диффузии, хотя оно и облегчается участием белка. Еще одним примером облегченной диффузии может служить перемещение хлорид- и гидрокарбонат-ионов между эритроцитами и плазмой крови при так называемом хлоридном сдвиге. Это один из механизмов, обеспечивающих частичную и избирательную проницаемость мембран.

Рис. 5.17. Облегченная диффузия с участием белка-переносчика. Белок пребывает попеременно в одном из двух состояний — «пинг» и «понг». Поскольку концентрация молекул глюкозы (шестиугольники) в наружной среде выше, реальный ее поток направлен в данном случае внутрь клетки — по диффузионному градиенту.
Осмос
Диффузия воды через полупроницаемые мембраны из области с высокой ее концентрацией в область с низкой концентрацией называется осмосом. Удобно рассматривать осмос как одну из форм диффузии, при которой перемещаются только молекулы воды. Обратимся к ситуации, представленной на рис. 5.18. Молекулы растворенного вещества в этом случае слишком велики, чтобы пройти через поры в мембране, так что равновесие может быть достигнуто только за счет перемещения молекул воды. В растворе А концентрация воды выше, поэтому реальный обусловленный осмосом поток воды направлен от А к В. По достижении равновесия реальный поток будет равен нулю. Стремление молекул воды перемещаться из одного места в другое измеряется водным потенциалом; обозначается эта величина греческой буквой — ψ («пси»). Вода всегда движется из области с высоким водным потенциалом в область с низким потенциалом.

Рис. 5.18. Два раствора, разделенных избирательно проницаемой мембраной.
Молекулы растворенного вещества снижают водный потенциал (в сущности, они «разбавляют» воду!). Степень этого снижения называют осмотическим потенциалом, ψо . Рис. 5.19 иллюстрирует влияние различных растворов на эритроциты. Об осмосе в растительных клетках мы будем говорить в гл. 13.

Рис. 5.19. Поведение эритроцитов в растворах разной концентрации. У гипотонического раствора водный потенциал выше, чем у содержимого эритроцита. Поэтому вода путем осмоса поступает в клетку и разрывает ее — содержимое эритроцита выходит наружу. У гипертонического раствора водный потенциал ниже, чем у клеточного содержимого и вода уходит из клетки — эритроцит сморщивается. В изотоническом растворе водные потенциалы раствора и клеточного содержимого равны, поэтому реального перемещения воды ни в ту, ни в другую сторону не происходит, и объем клетки не меняется, остается нормальным. Плазма крови должна быть изотоничной по отношению к эритроцитам и другим клеткам тела.
Активный транспорт
Активный транспорт — это сопряженный с потреблением энергии перенос молекул или ионов через мембрану против градиента концентрации. Энергия требуется потому, что вещество должно двигаться вопреки своему естественному стремлению диффундировать в противоположном направлении. Движение это обычно однонаправленное, тогда как диффузия обратима. Источником энергии для активного транспорта служит АТФ — соединение, образующееся в процессе дыхания и выполняющее в клетке роль носителя энергии. Поэтому в отсутствие дыхания активный транспорт идти не может.
Во внеклеточных и внутриклеточных жидкостях преобладают ионы натрия (Na + ), калия (K + ) и хлорид-ионы (Cl – ). На рис. 5.20 видно, что концентрации этих ионов внутри эритроцитов и в плазме крови человека весьма различны.

Рис. 5.20. Концентрация (в миллимолях) Na + , K + и Cl – в эритроцитах и в окружающей их среде.
Внутри эритроцитов, как и в большинстве клеток, концентрация калия значительно выше, чем снаружи. Другая характерная особенность заключается в том, что внутриклеточная концентрация калия превышает концентрацию натрия. Если каким-либо специфическим воздействием, например с помощью цианида, подавить дыхание эритроцитов, то их ионный состав начнет постепенно меняться и в конце концов сравняется с ионным составом плазмы крови. Это показывает, что данные ионы могут пассивно диффундировать через плазматическую мембрану эритроцитов, но что в норме за счет энергии, поставляемой процессом дыхания, идет их активный транспорт, благодаря которому и поддерживаются концентрации, указанные на рис. 5.20. Иными словами, натрий активно выкачивается из клетки, а калий активно накачивается в нее.
Активный транспорт осуществляется при помощи белков-переносчиков, локализующихся в плазматической мембране. Этим белкам в отличие от тех, о которых мы говорили при обсуждении облегченной диффузии, для изменения их конформации требуется энергия. Поставляет эту энергию АТФ, образующийся в процессе дыхания.
Сравнительно недавно выяснилось, что у большей части клеток в плазматической мембране действует натриевый насос, активно выкачивающий натрий из клетки. В животных клетках натриевый насос сопряжен с калиевым насосом, активно поглощающим ионы калия из внешней среды и переносящим их в клетку. Такой объединенный насос называют натрий-калиевым насосом [(Na + , K + )-насос]. Поскольку насос имеется почти во всех животных клетках и выполняет в них ряд важных функций, он представляет собой хороший пример механизма активного транспорта. О его физиологическом значении свидетельствует тот факт, что более трети АТФ, потребляемого животной клеткой в состоянии покоя, расходуется на перекачивание натрия и калия.

Рис. 5.21. Натрий-калиевый насос.
Насос — это особый белок-переносчик, локализующийся в мембране таким образом, что он пронизывает всю ее толщу (рис. 5.21). С внутренней стороны мембраны к нему поступают натрий и АТФ, а с наружной — калий. Перенос натрия и калия через мембрану совершается в результате конформационных изменений, которые претерпевает этот белок. Обратите внимание, что на каждые два поглощенных иона калия из клетки выводится три иона натрия. Вследствие этого содержимое клетки становится более отрицательным по отношению к внешней среде, и между двумя сторонами мембраны возникает разность потенциалов. Это ограничивает поступление в клетку отрицательно заряженных ионов (анионов), например хлорид-ионов.
Именно данным обстоятельством объясняется тот факт, что концентрация хлорид-ионов в эритроцитах ниже, чем в плазме крови (рис. 5.20), хотя эти ионы могут поступать в клетки и выходить из них за счет облегченной диффузии. Положительно заряженные ионы (катионы), напротив, притягиваются клеткой. Таким образом, оба фактора — концентрация и электрический заряд — важны при определении того, в каком направлении будут перемещаться через мембрану ионы.
Натрий-калиевый насос необходим животным клеткам для поддержания осмотического баланса (осморегуляции). Если он перестанет работать, клетка начнет набухать и в конце концов лопнет. Произойдет это потому, что с накоплением ионов натрия в клетку под действием осмотических сил будет поступать все больше и больше воды. Ясно, что бактериям, грибам и растениям с их жесткими клеточными стенками такой насос не требуется. Животным клеткам он нужен также для поддержания электрической активности в нервных и мышечных клетках и, наконец, для активного транспорта некоторых веществ, например сахаров и аминокислот. Высокие концентрации калия требуются также для белкового синтеза, гликолиза, фотосинтеза и для некоторых других жизненно важных процессов.
Активный транспорт осуществляется всеми клетками, но в некоторых случаях он играет особо важную роль. Именно так обстоит дело в клетках эпителия, выстилающего кишечник и почечные канальцы, поскольку функции этих клеток связаны с секрецией и всасыванием.
АКТИВНЫЙ ТРАНСПОРТ В КИШЕЧНИКЕ. Всасываясь в тонком кишечнике, продукты переваривания пищи должны пройти через клетки эпителия, выстилающего стенку кишки. Затем глюкоза, аминокислоты и соли через клетки, образующие стенки кровеносных сосудов, поступают в кровь и доставляются кровью в печень. Вскоре после приема пищи концентрация продуктов ее переваривания достигает в кишечнике довольно высокого уровня, так что всасывание в какой-то мере является и результатом диффузии. Однако диффузия здесь происходит очень медленно, и ее должен дополнять активный транспорт. Как видно из рис. 5.22, этот активный транспорт сопряжен с работой (Na + , K + )- насоса.
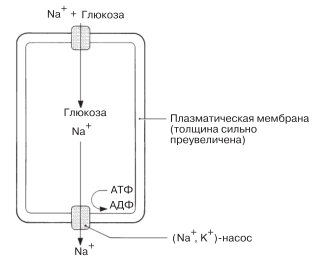
Рис. 5.22. Активный транспорт глюкозы через плазматическую мембрану клетки кишечника или почки. (На основе рис. 36.12 в книге L. Stryer (1981) Biochemistry, 2 nd ed., Freeman.)
Натрий, выкачиваемый из клетки натрий-калиевым насосом, стремится диффундировать обратно в клетку. В мембране находится транспортный белок, которому для выполнения его функций требуются натрий и глюкоза. Они транспортируются в клетку вместе пассивно, за счет облегченной диффузии. Активный транспорт аминокислот совершается при участии аналогичного белкового «натрий-аминокислотного» переносчика; активной частью этого процесса является выкачивание натрия наружу.
АКТИВНЫЙ ТРАНСПОРТ В НЕРВНЫХ И МЫШЕЧНЫХ КЛЕТКАХ. В нервных и мышечных клетках натрий-калиевый насос обеспечивает возникновение в плазматической мембране разности потенциалов, называемой потенциалом покоя (о проведении нервных импульсов см. гл. 17, а о мышечном сокращении — гл. 18). В мембранах саркоплазматического ретикулума мышечных клеток действует кальциевый насос; в этом случае в саркоплазматический ретикулум (специализированная форма эндоплазматического ретикулума) из окружающей его цитоплазмы активно накачивается кальций. Мышечное сокращение наступает в ответ на быстрое высвобождение кальция, вызванное нервным импульсом.
АКТИВНЫЙ ТРАНСПОРТ В ПОЧКАХ. В почках также имеет место активный транспорт: из проксимальных извитых канальцев почки активно транспортируются натрий и глюкоза, а в корковом веществе почки — натрий. Более подробно эти процессы рассматриваются в гл. 20.
АКТИВНЫЙ ТРАНСПОРТ В РАСТЕНИЯХ. Одним из примеров активного транспорта у растений может служить поступление сахаров во флоэму; по ней сахара доставляются в другие части растения (гл. 13). Особо важную роль играет этот процесс в листьях.
Эндоцитоз и экзоцитоз
Эндоцитоз и экзоцитоз — это два активных процесса, посредством которых различные материалы транспортируются через мембрану либо в клетки (эндоцитоз), либо из клеток (экзоцитоз) (рис. 5.23).

Рис. 5.23. Эндоцитоз и экзоцитоз.
При эндоцитозе плазматическая мембрана образует впячивания или выросты, которые затем, отшнуровываясь, превращаются в пузырьки или вакуоли. Различают два типа эндоцитоза.
- Фагоцитоз («поедание») — поглощение клетками твердых частиц. Специализированные клетки, осуществляющие фагоцитоз, называются фагоцитами; эту функцию выполняют, например, некоторые виды лейкоцитов, поглощающие бактерии. Мембранный мешочек, обволакивающий поглощаемую частицу, называют фагоцитозной вакуолью.
- Пиноцитоз («питье») — поглощение клеткой жидкого материала. Пузырьки, которые при этом образуются, часто бывают очень мелкими. В таком случае говорят о микропиноцитозеи пузырьки называют микропиноцитозными. Яйцеклетки человека именно таким способом поглощают питательные вещества из окружающих фолликулярных клеток.
В щитовидной железе гормон тироксин запасается в форме тиреоглобулина в особых полых структурах (фолликулах). Когда возникает потребность в тироксине, фолликулярные клетки поглощают тиреоглобулин путем пиноцитоза и здесь он превращается в тироксин, который затем поступает в кровь. Пиноцитоз характерен для очень многих клеток, как животных, так и растительных (рис. 5.10).
Экзоцитоз — процесс, обратный эндоцитозу. Таким способом различные материалы выводятся из клеток: из пищеварительных вакуолей удаляются оставшиеся непереваренными плотные частицы, а из секреторных клеток путем «пиноцитоза наоборот» выводится их секрет. Именно так секретируются в частности ферменты поджелудочной железы (рис. 5.29). В растительных клетках путем экзоцитоза экспортируются материалы, необходимые для построения клеточных стенок (рис. 5.30).