ФУНКЦИОНАЛЬНЫЕ ОТДЕЛЫ МОЗГА ЧЕЛОВЕКА
АНАТОМИЯ НЕРВНОЙ СИСТЕМЫ В.В. Жуков, Е.В. Пономарева – 1998
Спинной мозг
Спинной мозг находится внутри костного канала, образованного позвоночником, и удерживается в постоянном положении при помощи связок, идущих от мягкой и паутинной оболочек мозга к внутренней поверхности твердой оболочки. Последняя прикрепляется к позвонкам воронкообразными выростами в области межпозвоночных отверстий, через которые выходят корешки. Пространства между твердой, паутинной оболочками и мозгом (к которому плотно прилегает мягкая оболочка) заполнены спинномозговой жидкостью (СМЖ), которая также заполняет и центральный спинномозговой канал.
Характерная особенность организации спинного мозга — наличие правильной периодичности в выходе задних (дорсальных) и передних (вентральных) спинномозговых корешков, содержащих соответственно афферентные (центростремительные) и эфферентные (центробежные) волокна. На дорсальном корешке находится спинномозговой ганглий, содержащий афферентные нервные клетки, отростки которых входят в спинной мозг. Каждая пара корешков соответствует одному из позвонков и покидает позвоночный канал через отверстия между ними. После выхода из костного канала корешки сливаются и оба типа волокон смешиваются в нервах. Поэтому спинной мозг разделен на ряд сегментов (шейные, грудные, поясничные, крестцовые), число которых равно количеству позвонков. На третьем месяце внутриутробного развития плода спинной мозг занимает всю длину позвоночного канала. В последующие месяцы позвоночник растет в длину быстрее, чем спинной мозг, который у взрослых людей достигает 1-го поясничного позвонка. Каудальный конец спинного мозга имеет коническую форму (мозговой конус). Корешки нижних сегментов идут единым пучком, покрытым мозговой оболочкой (конский хвост). Отходящая от мозгового конуса тонкая фиброзная нить (концевая нить) прикрепляется к надкостнице крестца и фиксирует положение дистального конца спинного мозга. В поясничной и шейной области корешки образуют короткие смешанные нервы, формирующие поясничное и плечевое сплетения.
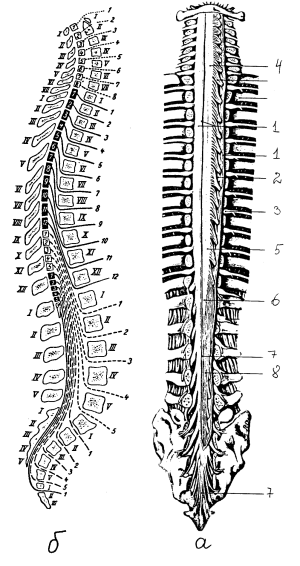
Рис. 7. Спинной мозг в позвоночном канале: а — внешний вид спинного мозга (твердая оболочка частично удалена): 1 — твердая оболочка и ее край, 2 — зубчатая связка, 3 – спинномозговой ганглий, 4 — шейное утолщение, 5 — поясничное утолщение, 6 — мозговой конус, 7 — концевая нить, 8 — конский хвост; б — схема расположения сегментов спинного мозга по отношению к позвоночному столбу: римские цифры — позвонки, арабские цифры — сегменты спинного мозга и спинномозговые нервы
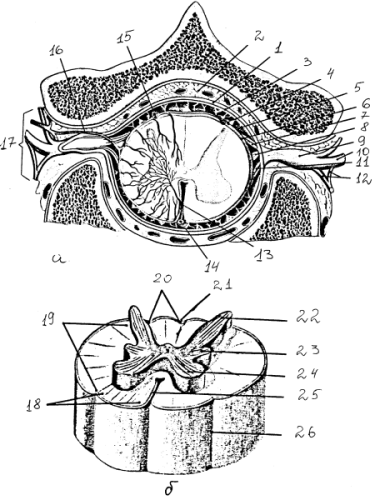
Рис. 8. Строение спинного мозга: а — поперечный срез через позвоночный канал на уровне 2-го грудного позвонка; б — белое и серое вещество спинного мозга. 1 — твердая оболочка, 2 — эпидуральное пространство, 3 – субдуральное пространство, 4 — паутинная оболочка, 5 — субарахноидальное пространство, 6 — мягкая оболочка, 7 — задний корешок, 8 — зубчатая связка, 9 — спинномозговой ганглий, 10 — спинной нерв, 11 — передний корешок, 12 – соединительные ветви, 13 — бороздчатокомиссуральная артерия, 14 — передняя спинномозговая артерия, 15 — задняя спинномозговая артерия, 16 — спинномозговая ветвь межреберной артерии, 17 — межпозвоночное отверстие, 18-20 — передний, боковой, задний канатики, 21 — задняя срединная борозда, 22-24 — задний, боковой, передний столбы серого вещества, образующие на поперечном сечении рога (задние, боковые, передние — соответственно), 25 – передняя срединная щель, 26 — передняя боковая борозда
В спинном мозге сосредоточены нервные клетки, аксоны которых дают начало нервам, идущим к поперечнополосатым мышцам тела. Эти и другие нейроны образуют серое вещество спинного мозга, расположенное вокруг его центрального канала. В нем различают передние и задние рога, а также разделяющую их промежуточную часть. Задние рога содержат нейроны, которые передают сенсорные импульсы к лежащим выше центрам или расположенным вентральнее двигательным клеткам, замыкая рефлекторные дуги. В передних рогах находятся самые крупные нейроны, иннервирующие мышцы (двигательные нейроны, или мотонейроны). В промежуточной зоне находятся клетки серого вещества, связывающие его чувствительный и двигательный отделы. С 1-го грудного сегмента спинного мозга до 3-го поясничного в боковых отделах серого вещества расположены боковые столбы, состоящие из двигательных нейронов симпатической части автономной нервной системы. Расположение нейронов в сером веществе неравномерно: определенные их типы собираются в группы (столбы, или ядра).
Серое вещество спинного мозга окружено слоем белого вещества, состоящего из восходящих и нисходящих миелинизированных волокон. Белое вещество разделяется выростами серого на каждой стороне на три канатика: передний (вентральный), задний (дорсальный) и боковой (латеральный). Восходящие пути передают сенсорную информацию к рефлекторным подкорковым центрам и коре головного мозга, нисходящие проводят информацию к двигательным клеткам передних рогов. Системы коротких волокон (проприоспинальные пути) связывают между собой разные сегментарные уровни спинного мозга.
Проводящие пути
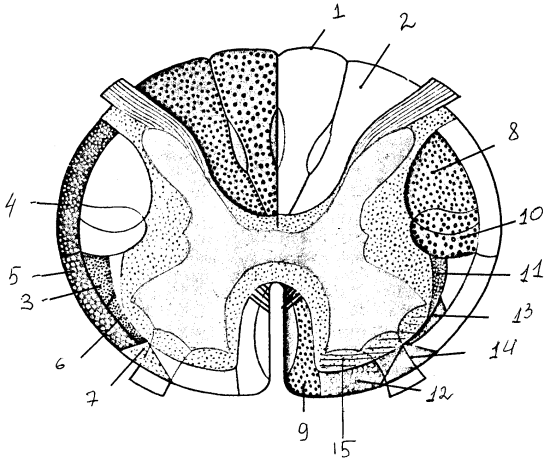
Рис. 9. Проводящие пути спинного мозга: восходящие пути: 1 — тонкий пучок, 2 — клиновидный пучок, 3 — спинно-таламический путь, 4 — спинно-мозжечковый задний путь, 5 — спинно-мозжечковый передний путь, 6 — спинно-тектальный путь, 7 — спинно-оливарный путь; нисходящие пути: 8 — кортико-спинальный латеральный (пирамидный) путь, 9 — кортико-спинальный передний (пирамидный) путь, 10 — рубро-спинальный путь, 11, 12 — вестибуло-спинальные (латеральный и передний) пути, 13 — текто-спинальный путь, 14 — оливо-спинальный путь, 15 — ретикуло-спинальный путь.
Восходящие пути
1. Тонкий пучок (Голля) и клиновидный пучок (Бурдаха) проходят в дорсальных канатиках и образованы аксонами афферентных неронов спинномозговых ганглиев. Нейроны 2-го порядка находятся в продолговатом мозге в ядрах Голля (медиальные) и Бурдаха (латеральные). Аксоны нейронов этих ядер в стволе мозга совершают перекрест (медиальная петля). Нейроны 3-го порядка находятся в ядрах промежуточного мозга и направляют аксоны в кору больших полушарий. По этим путям в ЦНС поступает сенсорная информация главным образом от проприоцепторов мышц и сухожилий, а также от тактильных рецепторов кожи.
2. Спинно-таламический путь обеспечивает передачу в основном болевой и температурной чувствительности. Нейроны 1-го порядка располагаются в спинномозговых ганглиях, а 2-го порядка — в основании дорсальных рогов серого вещества спинного мозга. Аксоны последних переходят на противоположную сторону и по белому веществу латерального или вентрального канатика проходят без перерыва до промежуточного мозга. Расположенные в нем нейроны 3-го порядка посылают аксоны в кору больших полушарий.
3. Спинно-мозжечковые пути — дорсальный (пучок Флексига) и вентральный (пучок Говерса) — занимают латеральные канатики. Нейроны 1-го порядка также находятся в спинальных ганглиях. Аксоны нейронов 2-го порядка, расположенные в сером веществе спинного мозга, идут по латеральным канатикам той же (путь Флексига) или противоположной (путь Говерса) стороны к мозжечку. В стволе мозга волокна пути Говерса снова перекрещиваются и поступают в кору мозжечка той же стороны. Оба пути обеспечивают передачу сигналов от проприоцепторов мышц и сухожилий, а также висцерорецепторов.
Таким образом, кора больших полушарий связана с афферентными волокнами противоположной стороны тела, а кора мозжечка связана преимущественно с нейронными структурами одноименной стороны.
Нисходящие пути
1. Кортико-спинальный (пирамидный) путь образован аксонами пирамидных клеток V слоя преимущественно двигательных областей коры больших полушарий. Эти аксоны проходят без перерыва через ствол мозга. Большая часть их в области продолговатого мозга переходит на противоположную сторону (перекрещенный путь) и проходит в вентральном канатике до соответствующих сегментов спинного мозга. Небольшая часть волокон не претерпевает перекреста в продолговатом мозге, однако и они переходят на противоположную сторону в сегментах спинного. В обоих случаях двигательная кора больших полушарий оказывается напрямую связанной с нейронами противоположной стороны спинного мозга. Этот путь филогенетически один из самых молодых в ЦНС и развит только у млекопитающих. Он связывает пирамидные клетки двигательной коры, как правило, с интернейронами (у приматов также и с мотонейронами).
2. Рубро-спинальный путь образован аксонами нейронов красного ядра среднего мозга. Эти аксоны перекрещиваются непосредственно под ядром и по латеральному канатику спускаются до различных сегментов спинного мозга, устанавливая в его сером веществе связи с интернейронами. Это более древняя, чем пирамидный путь, система контроля двигательной функции и развита не только у млекопитающих, но и у рептилий и птиц.
3. Вестибуло-спинальный путь образован нейронами расположенных в продолговатом мозгу ядер Дейтерса. Их аксоны проходят в вентральном канатике и заканчиваются на интер- и мотонейронах промежуточной зоны серого вещества спинного мозга.
4. Ретикуло-спинальный путь образован аксонами ретикулярной формации заднего мозга. Основная часть этих волокон заканчивается на интернейронах медиальной части вентрального рога.
Задний мозг (ромбовидный мозг)
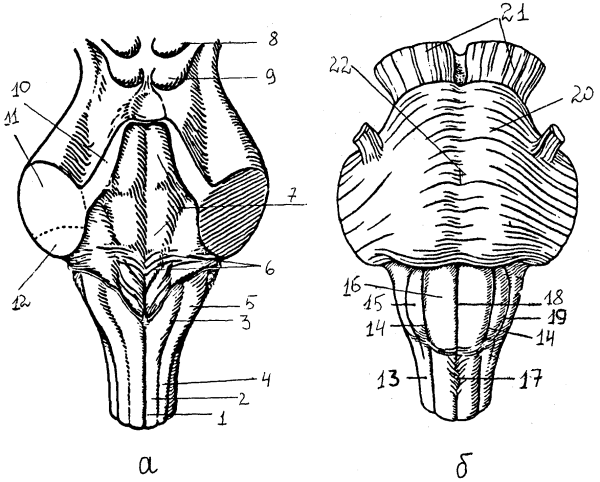
Рис. 10. Ствол мозга: а — задняя поверхность (мозжечок удален): 1 — задняя срединная борозда, 2 — тонкий пучок, 3 — бугорок тонкого ядра, 4 — клиновидный пучок, 5 — бугорок клиновидного ядра, 6 — мозговые полоски, 7 — ромбовидная ямка, дно IY желудочка, 8, 9 — верхние и нижние бугорки четверохолмия, 10-12 — верхняя, средняя, нижняя мозжечковые ножки; б — передняя поверхность: 13 — боковой канатик, 14 — передне-латеральная борозда, 15 — олива, 16 — пирамида, 17 — перекрест пирамид, 18 – передняя срединная щель, 19 — позади-оливная борозда, 20 — мост, 21 — ножки мозга, 22 — базилярная борозда
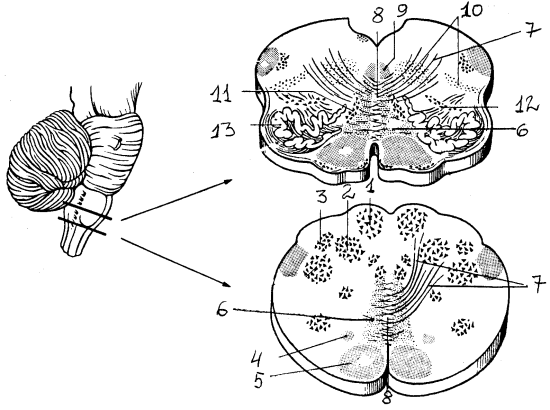
Рис. 11. Поперечный разрез продолговатого мозга (на двух уровнях): 1 — тонкое ядро, 2 — клиновидное ядро, 3 — добавочное клиновидное ядро, 4 — текто-спинальный путь, 5 — пирамидный пучок, 6 — медиальная петля, 7 — внутренние дугообразные волокна, начинающиеся от тонкого и клиновидного ядер, формируют медиальную петлю, 8 — перекрест медиальных петель, 9 — медиальный продолговатый пучок, 10 — ретикулярная формация, 11, 12 — медиальное и заднее добавочные оливные ядра, 13 — нижнее оливное ядро
Задний мозг включает в себя продолговатый мозг и варолиев мост и сохраняет черты сегментарного строения, что проявляется в расположении ядер V — XII пар черепно-мозговых нервов. Характерной особенностью является наличие мощной массы нервных элементов, которые не получают непосредственно афферентных волокон из периферических источников и не отдают двигательных волокон, — ретикулярной формации. Эта надсегментарная структура занимает всю центральную часть заднего мозга и доступна для вмешательства со стороны дна IV желудочка. Другие надсегментарные структуры — ядра Голля и Бурдаха, а также расположенные в толще мозга оливы.
Средний мозг
Средний мозг состоит из трех основных частей: крыша, покрышка и ножки мозга. Крыша образована парами верхних и нижних бугров четверохолмия. Между крышей и расположенной вентральнее покрышкой проходит сильвиев водопровод. В покрышке находятся ядра III и IV пар черепномозговых нервов, а также добавочное ядро III нерва (парасимпатическое ядро Эдингера — Вестфаля), парные красные ядра и несколько ядерных скоплений меньшего размера, относящихся к ретикулярной формации. Ножки мозга содержат главным образом нисходящие двигательные волокна, начинающиеся в коре мозга и идущие к ядрам черепномозговых нервов и двигательным ядрам спинного мозга. Непосредственно под красными ядрами находится группа клеток, составляющих черное вещество, анатомически входящее в состав ножек мозга.
Мозжечок
Основные части мозжечка — два полушария и непарный червь. Со стволовой частью мозжечок соединяется тремя парами ножек: верхними — со средним мозгом, средними — с варолиевым мостом, нижними — с задним мозгом. Его поверхность разделяется глубокими бороздами на доли и извилины. По Ларселу, поверхность мозжечка делят на отделы в зависимости от поступающих афферентных путей и филогенетического возраста. Филогенетически более древний архицеребеллум занимает наиболее каудальную часть (флоккуло-нодулярная доля). Палеоцеребеллум включает в себя переднюю часть червя, пирамиду, язычок и парафлоккулярный отдел. Масса мозжечка состоит из покрывающей его поверхность коры и расположенного глубже белого вещества с ядрами.
Кора состоит из трех слоев.
1. Поверхностный молекулярный состоит из параллельных волокон и разветвлений дендритов нижележащих клеток. В нижней части расположены тела корзинчатых клеток, аксоны которых оплетают клетки Пуркинье, а также звездчатые клетки.
2. Ганглиозный слой — тела клеток Пуркинье. Эти клетки имеют обильно разветвленное дендритное дерево, которое направляется к поверхности мозжечка, и аксон, идущий в белое вещество. Аксоны клеток Пуркинье — единственные выход из коры мозжечка, они заканчиваются на нейронах ядер мозжечка и вестибулярных ядер.
3. Гранулярный слой содержит большое число тел клеток-зерен. Их аксоны поднимаются вертикально в молекулярный слой и там Т-образно ветвятся, образуя параллельные волокна, которые формируют синапсы на дендритах других клеток. В этом же слое находятся клетки Гольджи, аксоны которых подходят к клеткам-зернам.
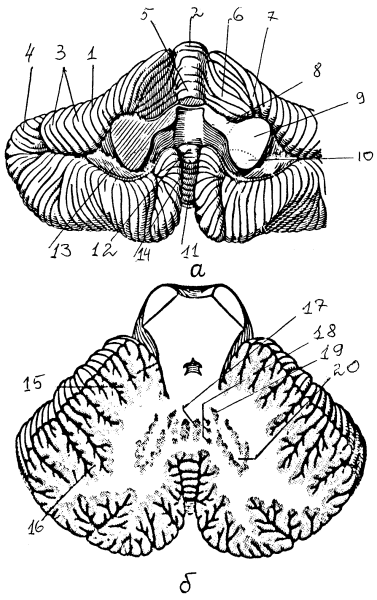
Рис. 13. Строение мозжечка: а — вид снизу, б — горизонтальный разрез: 1 — полушарие мозжечка, 2 — вершина, 3 — листки мозжечка, 4 — борозда, 5 — язычок, 6 — крыло центральной дольки, 7 — четырехугольная долька (передняя часть), 8-10 — верхняя, средняя и нижняя мозжечковые ножки, 11 — пирамида, 12 — узелок, 13 — клочок, 14 — язычок червя, 15 — кора мозжечка, 16 — древо жизни мозжечка, 17 — ядро шатра, 18 — шаровидное ядро, 19 — пробковидное ядро, 20 — зубчатое ядро
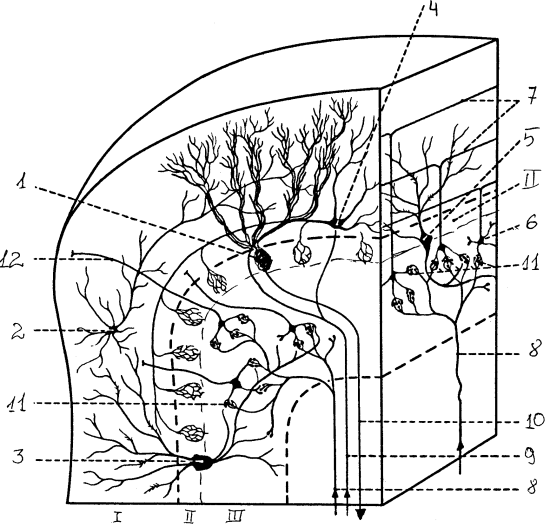
Рис. 14. Кора мозжечка и ее нейронный состав: I — молекулярный слой, II — ганглиозный слой, Ш — гранулярный слой; 1 — клетка Пуркинье, 2 — звездчатая клетка, 3 — клетка Гольджи, 4 — корзинчатая клетка, 5 — клетка Гольджи, 6 — клетка-зерно, 7 – параллельные волокна, 8 — мшистые волокна, 9 — лазающие волокна, 10 — аксон клетки Пуркинье, 11 — гломерулы мозжечка, 12 — аксон зернистой клетки
Афферентный вход мозжечка образован тремя системами волокон. Во-первых, идущие из олив продолговатого мозга лазающие (лиановидные) волокна, заканчивающиеся синапсами на дендритах клеток Пуркинье. Во-вторых, мшистые (моховидные) волокна идут от ядер моста и заканчиваются на клетках-зернах. В-третьих, адренэргические волокна из голубого пятна, лежащего на границе заднего и среднего мозга.
Подкорковые парные ядра мозжечка: ядро шатра, шаровидное, пробковидное и зубчатое. Аксоны нейронов этих ядер идут к стволовым структурам: от ядер шатра — к вестибулярному ядру Дейтерса и ретикулярной формации, от шаровидных и пробковидных — к красным ядрам, от зубчатых — к таламусу и далее к коре больших полушарий.
Афферентные связи мозжечка различны в его отделах: архицеребеллум получает связи по вестибуло-мозжечковому пути, палеоцеребеллум — по спинно-мозжечковому пути, а неоцеребеллум — также по спинно-мозжечковым путям, по корково-мостовым путям, главным образом от сенсо-моторной коры больших полушарий.
Промежуточный мозг
Клеточные скопления промежуточного мозга локализуются вокруг третьего желудочка, образуя его боковые, верхние и нижние стенки. Они могут быть поделены на 5 отделов: зрительный бугор (таламус), забугорная область (метаталамус), подбугорная область (гипоталамус), надбугорная область (эпиталамус), субталамическая область (субталамус).
В средней части обе половины таламуса соединяются между собой вместе перемычкой — промежуточной массой. Анатомически ядра таламуса делят на группы: передние, интраламинарные, срединные, боковые и задние. Ядра, дающие четко очерченные проекции в сенсорную кору больших полушарий, называют специфическими, или проекционными. В них заканчиваются длинные аксоны нейронов восходящих афферентных систем (кроме обонятельного пути). Неспецифические ядра связаны с ретикулярной формацией и дают в коре полушарий диффузные проекции. Выделяют также ассоциативные ядра, получающие сигналы от специфических, и моторные ядра, включенные в систему регуляции движений.
К метаталамусу относятся латеральные (ЛКТ) и медиальные (МКТ) коленчатые тела, являющиеся проекционными ядрами.
В состав эпиталамуса входит шишковидная железа (эпифиз) и ядра уздечки.
К субталамусу относится ядерное образование с тем же названием.
В гипоталамус входят: перекрест зрительных нервов, заканчивающаяся гипофизом воронка, серый бугор и наиболее каудально расположенные сосцевидные тела.
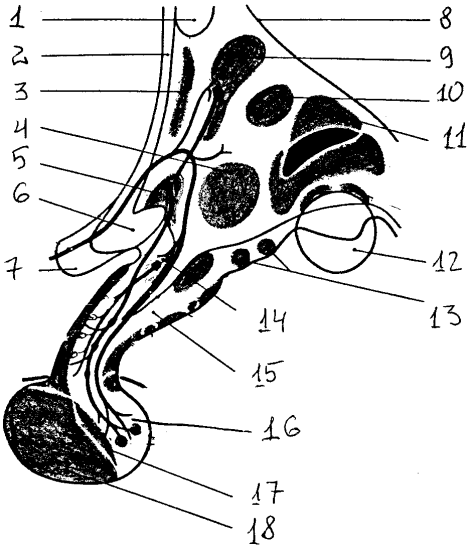
Рис. 16. Ядра гипоталамуса : 1 — передняя комиссура, 2 — конечная пластинка, 3 – преоптическая область, 4 — вентромедиальное ядро, 5 — супраоптическое ядро, 6 — зрительный перекрест, 7 — зрительный нерв, 8 — гипоталамическая борозда, 9 — паравентрикулярное ядро, 10 — дорсомедиальное ядро, 11 — заднее ядро, 12 — сосцевидное тело, 13 — латеральные ядра серого бугра, 14 — ядра воронки, 15 — воронка, 16 — нейрогипофиз, 17 — промежуточная доля, 18 — передняя доля
В этих образованиях расположено большое количество ядер, которые можно разделить на несколько групп: преоптическую, переднюю, среднюю, наружную и заднюю. Наиболее хорошо выделяются паравентрикулярное и супраоптическое ядра (передняя группа), заднее гипоталамическое и мамиллярное ядра (задняя группа). Среди сложных и запутанных связей гипоталамуса можно выделить короткий, но очень мощный пучок волокон (до 100 000 аксонов), заканчивающийся в задней доле гипофиза. По этим волокнам происходит не только распространение нервных волокон, но и ток нейросекрета, вырабатываемого клетками супраоптического и паравентрикулярных ядер. Гипофиз, являющийся важнейшей железой внутренней секреции, состоит из трех долей: передней (аденогипофиз), промежуточной и задней (нейрогипофиз) — из которых только последняя является производным нервной системы.
Таблица 1
Черепномозговые нервы и их функции
| Номер пары | Наименование нерва | Функция |
| I | Обонятельный | Сенсорный вход от обонятельного эпителия |
| II | Зрительный | Сенсорный вход от ганглиозных клеток сетчатки |
| III | Глазодвигательный | Моторный выход к четырем из шести наружных мышц глазного яблока |
| IV | Блоковый | Моторный выход к верхней косой мышце глазного яблока |
| V | Тройничный | Основной сенсорный вход от лица. Моторный выход к жевательным мышцам |
| VI | Отводящий | Моторный выход к наружной прямой мышце глазного яблока |
| VII | Лицевой | Основной моторный выход к мышцам лица. Сенсорный вход от некоторых вкусовых рецепторов |
| VIII | Слуховой | Сенсорный вход от внутреннего уха и вестибулярного органа |
| IX | Языкоглоточный | Сенсорный вход от рецепторов (в том числе вкусовых) языка и глотки |
| X | Блуждающий | Главный парасимпатический моторный выход к мышцам многих внутренних органов: сердца, желудка, кишечника и др. Моторный выход к мышцам глотки. Сенсорный вход от некоторых вкусовых рецепторов |
| XI | Добавочный | Моторный выход к грудино-ключично-сосковой и трапециевидной мышцам |
| XII | Подъязычный | Моторный выход к мышцам языка |
Конечный мозг
Конечный мозг состоит из покрытых корой двух полушарий и базальных (подкорковых) ядер. У взрослого человека поверхность полушарий делится на лобную, теменную, затылочную и височную доли. Каждый из этих отделов разделяется бороздами на меньшие по размерам извилины: верхние, средние и нижние височные и лобные, прецентральную и постцентральную и т.д. Латеральная (сильвиева) борозда отделяет височную долю от теменной и лобной, границу между которыми образует центральная (роландова) борозда. Отдельно расположен островок, лежащий в глубине латеральной борозды на боковой поверхности полушария и прикрытый лобной, височной и теменной долями. Выделяют также лимбическую долю, в которую входят части лобной, теменной и височной долей: подмозолистая область, поясная извилина, перешеек поясной извилины, парагиппокампальная извилина, крючок гиппокампа, миндалина и диагональная связка Брока.
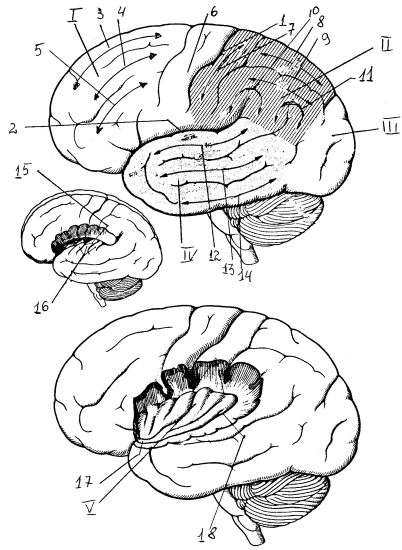
Рис. 17. Латеральная поверхность полушарий: I — Y — лобная, теменная, затылочная, височная и островковая доли; 1 — центральная борозда, 2 — латеральная борозда, 3, 4, 5 — верхняя, средняя, нижняя лобные извилины, 6, 7 — прецентральная и постцентральная извилины, 8, 9 — верхняя, нижняя теменные дольки, 10 — надкраевая извилина, 11 – угловая извилина, 12, 13, 14 — верхняя, средняя, нижняя височные извилины, 15 — поперечные височные борозды, 16 — поперечные височные извилины (Гешля), 17 — порог островка, 18 — круговая борозда островка
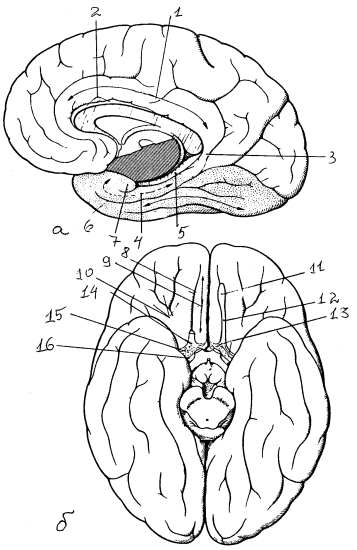
Рис. 18. Полушария большого мозга: а — медиальная поверхность, б — основание полушарий: 1 — поясная извилина, 2 — борозда мозолистого тела, 3 — перешеек поясной извилины, 4 — парагиппокампальная извилина, 5 — зубчатая извилина, 6 — носовая борозда, 7 — крючок, 8 — обонятельная борозда, 9 — прямая извилина,10 — глазничные извилины, 11 — обонятельная луковица, 12 – обонятельный тракт, 13 — медиальная и латеральная обонятельные полоски, 14 – обонятельный треугольник, 15 — переднее продырявленное вещество, 16 – диагональная извилина Брока, в глубине которой проходит компактный пучок волокон — диагональная связка Брока
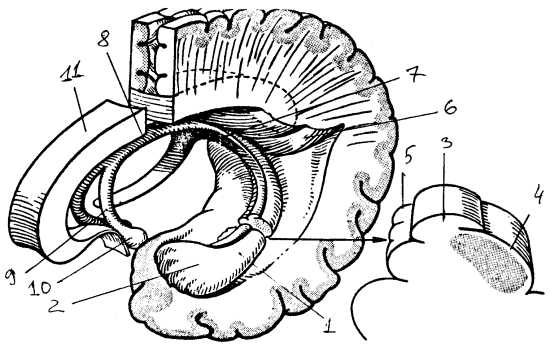
Рис. 19. Гиппокамп левого полушария: 1 — гиппокамп, 2 — ножка, 3 — бахромка, 4 — лоток, 5 — зубчатая извилина, 6 — птичья шпора, 7 — ножка свода, 8 — тело свода, 9 -столб, 10 — сосцевидное тело, 11 — мозолистое тело

Рис. 20. Базальные ядра на горизонтальном (а) и фронтальном (б) разрезах полушарий головного мозга: 1, 2 — головка и тело хвостатого ядра, 3 — ограда, 4 — скорлупа, 5, 6 — латеральный и медиальный сегменты бледного шара, 7,8 – наружная и медиальная мозговые пластинки, 9-11 — самая наружная, наружная и внутренняя капсулы, 12 — таламус
Части коры млекопитающих содержат следы филогенетического развития. Палеокортекс: обонятельные луковицы (в которые непосредственно поступают волокна от обонятельного эпителия), обонятельные тракты (расположены на нижней поверхности лобных долей), обонятельные бугорки (вторичные обонятельные центры) и некоторые другие близлежащие структуры.
Архекортекс: поясная извилина, извилина морского коня (извилина гиппокампа), миндалевидное тело (скопление серого вещества под корой височной доли).
Остальные части коры относят к неокортексу, который у млекопитающих занимает доминирующее положение, оттесняя старую кору вперед и внутрь.
Комплекс базальных ядер (полосатое тело) расположен в толще конечного мозга и состоит из хвостатого ядра, бледного шара, скорлупы и ограды. Полоска белого вещества (внутренняя капсула), проходящая между таламусом и хвостатым ядром с одной стороны и бледным шаром — с другой. Наружная капсула отделяет полосатое тело от ограды.
Основной структурной особенностью коры является правильная ориентация клеточных элементов и волокон, которые идут в направлении перпендикулярном поверхности, либо параллельно ей. Обычно близкие по размеру клетки группируются на одинаковой глубине, образуя клеточные слои.
Типичной для млекопитающих является шестислойная кора: 1) верхний молекулярный (плексиморфный) слой содержит мелкие нервные клетки и апикальные волокна пирамидных клеток нижележащих слоев; 2) наружный зернистый слой содержит пирамидные клетки небольшого размера; 3) слой средних пирамид; 4) внутренний зернистый слой состоит из большого количества короткоаксонных нейронов; 5) внутренний пирамидный слой содержит особенно большие пирамидные нейроны; 6) полиморфный слой. Однако в различных участках коры имеются определенные различия в нейронной организации, что лежит в основе создания карт цитоархитектонического строения коры. Например, на карте Бродмана выделено 11 областей, включающих 52 поля.
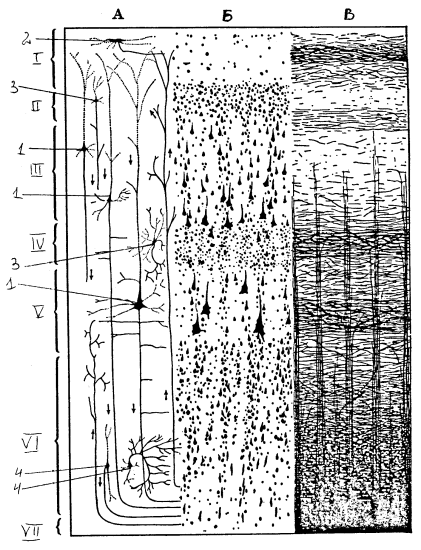
Рис. 21. Строение коры больших полушарий: А — слои коры, Б — цитоархитектоника, В — миелоархитектоника; слои: I — молекулярный, II — наружный зернистый, III – внешний слой пирамидных клеток (средних пирамид), IY — внутренний зернистый, Y — внутренний пирамидный, YI — полиморфный, YII — белое вещество; 1 — пирамидные клетки разных размеров, 2 — клетка Кахаля, 3 — звездчатые клетки, 4 — непирамидные клетки различного типа