Физиология поджелудочной железы
- Эндокринная функция
- Экзокринная функция
- Секреция жидкости и электролитов
- Синтез и секреция ферментов. Амилаза, липаза, протеазы
- Стимуляторы панкреатической секреции
- Фазы пищеварения
- Ингибиторы секреции поджелудочной железы
Эндокринная функция
Выделяют четыре типа эндокринных клеток. В-клетки (Р) наиболее многочисленны в островках Лангерганса, они секретируют инсулин и локализуются в центре островков. Другие эндокринные клетки расположены по периферии островков вокруг В-клеток: А-клетки (а), секретирующие глюкагон; D-клетки, секретирующие соматостатин; F-клетки (РР), продуцирующие панкреатический полипептид. Соотношение А-, D- и F-клеток, расположенных по периферии, неодинаково в каждом ацинусе. В передней части железы больше встречается F-клеток, тогда как в задней части железы содержится больше А-клеток. Физиологическое значение такой региональной вариации до конца не изучено, но наличие клеток разных типов необходимо для паракринной регуляции функции островков с помощью одного гормона, соматостатина, отвечающего за высвобождение других «гормонов островков» — инсулина и глюкагона.
Экзокринная функция
Экзокринная секреция поджелудочной железы состоит в выделении пищеварительных ферментов и жидкости, богатой электролитами. Как уже упоминалось, ацинарные клетки отвечают за синтез и секрецию пищеварительных ферментов, а центроацинарные клетки и эпителиальные клетки протоков — за секрецию жидкости, которая транспортирует ферменты в двенадцатиперстную кишку, где они активируются. Каждый из этих процессов подробно обсуждается далее.
Секреция жидкости и электролитов
Гастроинтестинальный гормон секретин стимулирует секрецию воды, бикарбонатов, натрия, калия и хлоридов эпителием протоков посредством активации аденилатциклазы. Последующее образование циклического аденозинмонофосфата (цАМФ) стимулирует хлорные каналы на люминальной стороне эпителиальных клеток, после чего хлориды высвобождаются из цитоплазмы в просвет протоков. Механизм хлоридно-бикарбонатного обмена способствует обмену хлоридов на внутриклеточные бикарбонаты, и, таким образом, продуцируется богатая бикарбонатами жидкость, необходимая для транспорта пищеварительных ферментов. Холинергическая стимуляция может вызывать сходный эффект, независимо от действия секретина. Поэтому при холинергической стимуляции секрет железы богат бикарбонатами и беден хлоридами, а в состоянии покоя возникает противоположная ситуация. В покое жидкость протоков секретируется со скоростью около 0.2 мл/мин, тогда как в процессе стимуляции приближается к 4 мл/мин (рис. 8.11). Всего поджелудочная железа секретирует в двенадцатиперстную кишку около 2.5 л жидкости каждые сутки.
Синтез и секреция ферментов
Ферменты поджелудочной железы образуются и хранятся в ацинарных клетках. В базальной части клетки расположены ядро и шероховатый эндоплазматический ретикулум, в котором происходит синтез белка. Ферменты из шероховатого эндоплазматического ретикулума поступают в комплекс Гольджи, находящийся между ядром и апикальной частью клетки, где они упаковываются в зимогенные гранулы и хранятся (рис. 8.12) до момента стимуляции клетки. После стимуляции, например пищей, отмечается уменьшение гранул в размерах и их числа в клетках. Соответственно, результатом этого является увеличение секреции ферментов поджелудочной железы. Каждая зимогенная гранула содержит в различном соотношении все ферменты поджелудочной железы. Ферменты в гранулах обычно находятся в «уплотненном» состоянии и растворяются после их экскреции из клетки в щелочной секрет поджелудочной железы. Однако растворение ферментов происходит в неактивной (проферментной) форме, а переход в активную форму осуществляется не ранее, чем они попадут в двенадцатиперстную кишку. В этом заключается механизм защиты поджелудочной железы от самопереваривания. Кроме того, зона плотного соединение межклеточных контактов апикальных частей клеток поджелудочной железы препятствует рефлюксу пищеварительных ферментов из просвета протоков в межклеточное пространство и служит еще одним механизмом защиты поджелудочной железы. При попадании в двенадцатиперстную кишку кислоточувствительные ферменты поджелудочной железы защищены от кислотного расщепления секретом железы, имеющим щелочную среду, в котором они были транспортированы. Эти предшественники ферментов активируются посредством ферментативного гидролиза, о котором будет рассказано ниже.
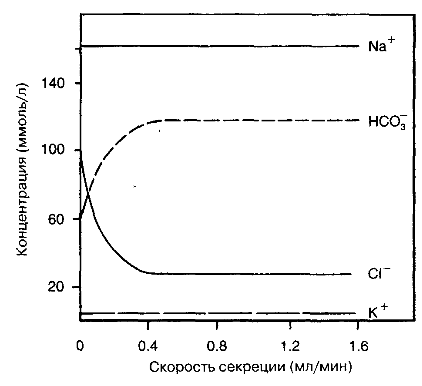
Рис. 8.11. Отношение содержания электролитов в секрете поджелудочной железы к скорости секреции.

Рис. 8.12. Строение ацинуса поджелудочной железы. Показано расположение зимогенных гранул но отношению к просвету протока.
Поджелудочная железа секретирует большое количество пищеварительных ферментов (табл. 8.1). Большинство из них предназначены для переваривания белков, жиров, углеводов, потребляемых с пищей. Для того, чтобы ферменты начали функционировать, они должны быть активированы в двенадцатиперстной кишке. Профермент трипсиноген подвергается ферментативному гидролизу с N-терминального фрагмента благодаря активности пептидазы (энтерокиназы), располагающейся на щеточной кайме энтероцитов тонкой кишки. Щеточная каемка тонкой кишки состоит из ворсинок, микроворсинок и крипт. В дополнение к механизмам, обеспечивающим абсорбцию питательных веществ, клетки щеточного барьера кишки выделяют различные вещества, способствующие пищеварению до момента абсорбции. Энтерокиназа является одним из таких веществ. Активированный трипсин, в свою очередь, катализирует активацию других проферментов, секретируемых поджелудочной железой. Поджелудочная железа также секретирует ингибитор трипсина. Этот пептид инактивирует трипсин, соединяясь с ним около его каталитического центра, и также является механизмом защиты поджелудочной железы. Механизм обратной связи регуляции процессов пищеварения с участием двенадцатиперстной кишки будет рассмотрен ниже.
Амилаза
Амилаза секретируется не только поджелудочной железой, но и слюнными железами. Несмотря на то, что две изоформы фермента имеют одинаковую энзимати-ческую активность, они могут быть разделены по своей электрофоретической подвижности. Амилаза участвует в расщеплении крахмала (углевод растительного происхождения) и гликогена (углевод животного происхождения). Амилаза слюнных желез начинает этот процесс и может фактически совершить переваривание значительной части крахмала до поступления его в тонкую кишку и контакта с панкреатической амилазой. Амилаза гидролизует а 1,4 гликозидные связи крахмала и гликогена, но не способна расщеплять а l ,6 связи. Продуктами переваривания амилазой крахмала являются полисахариды — а-декстрины с а1,6 связями, а l,6 связи гидролизуются другими кишечными ферментами после разрыва а1,4 связей. Таким образом, в процессе действия амилазы образуются вещества с а1,4 связями — мальтоза и мальтотриоза. Эти сахара разрушаются ферментами интестинальной щеточной каемки и обеспечивают вход глюкозы в эпителиальные клетки тонкой кишки.
Таблица 8.1. Пищеварительные ферменты поджелудочной железы
| Фермент | Мишень |
| Амилаза | а1,4 гликозидные связи крахмала, гликогена |
| Липаза | Триглицериды (образование 2-моноглицеридов и жирных кислот) |
| Фосфолипаза А2 | Фосфатидилхолин (образование лизофосфатидилхоли-на и жирных кислот) |
| Карбоксилэстераза | Эфиры холестерина, эфиры жирорастворимых витаминов; три-, ди-, моноглицериды |
| Трипсин* | Внутренние связи белка (основные аминокислоты) |
| Химотрипсин* | Внутренние связи белка (ароматические аминокислоты, лейцин, глутамин, метионин) |
| Эластаза* | Внутренние связи белков (нейтральные аминокислоты) |
| Карбоксипептидаза А и В* | Наружные связи белков, включая ароматические и нейтральные алифатические аминокислоты (А) и основные аминокислоты (В) с карбоксильного конца |
*Эти ферменты секретируются поджелудочной железой в неактивной форме (проферменты). Они активируются в двенадцатиперстной кишке
Липаза
Панкреатическая липаза катализирует расщепление триглицеридов пищи до двух жирных кислот и моноглицерида. Хотя липаза имеет некоторую самостоятельную активность, основное свое действие она осуществляет вместе с желчными кислотами, секретируемыми печенью, и колипазой поджелудочной железы, которая требуется для проявления полной активности липазы.
Желчные кислоты действуют как эмульгатор, формируя мелкие частицы жира и создавая условия для лучшего доступа липазы. Колипаза, липаза и соли желчных кислот формируют комплекс, с помощью которого увеличивается площадь поверхности действия липазы. Поджелудочная железа секретирует две формы липазы: фосфолипазу Af, расщепляющую фосфатидилхолин до лизофосфатидилхолина и свободной жирной кислоты, и карбоксилэстеразу, действующую на различные субстраты, включая эфиры холестерина, три-, ди-, и моноглицериды и эфиры жирорастворимых витаминов.
Протеазы
Поджелудочная железа секретирует различные протеазы в виде форм-предшественников, которые активируются в двенадцатиперстной кишке. Трипсин, химотрипсин и эластаза являются эндопептидазами, которые расщепляют белки в местах соединения специфических аминокислот. Карбоксипептидазы расщепляют связи пептидов на карбокси-терминальных окончаниях белков. В результате комбинированной активности этих эндопептидаз и карбоксипептидаз образуются олигопептиды и некоторые свободные аминокислоты, а олигопептиды в дальнейшем расщепляются ферментами щеточной каемки или поступают в клетки слизистой оболочки тонкой кишки.
Регуляция секреции
Большинство регуляторов секреции ферментов поджелудочной железы действуют на рецепторы мембраны ацинарных клеток, располагающиеся на базолатеральной поверхности этих клеток. Выделяют рецепторы для холецистокинина, бомбезина, ацетилхолина, субстанции Р, вазоактивного интестинального пептида (ВИП), секретина. Некоторые из этих веществ оказывают стимулирующее действие, другие — ингибирующее.
Стимуляторы панкреатической секреции
ВИП и секретин стимулируют панкреатическую секрецию, активируя аденилатциклазу. Как и в других типах клеток, аденилатциклаза способствует образованию цАМФ, в результате чего активируется протеинкиназа А, которая усиливает секрецию панкреатического сока, богатого бикарбонатами.
Другие агонисты (холецистокинин, ацетилхолин, гастрин-рилизинг пептид, субстанция Р) действуют через специфические рецепторы, в которых альтернативные «вторичные мессенджеры» задействованы в большей степени, чем цАМФ. Эти вещества повышают внутриклеточное содержание цГМФ, что приводит к увеличению внутриклеточного содержания инозитолтрифосфата, диацилглицерола, арахидоновой кислоты и кальция (рис. 8.13). Эти промежуточные вещества-посредники активируют различные протеинкиназы, результатом чего является повышение секреции ферментов. Данные, полученные в опытах на животных, свидетельствуют, что действие комбинации агонистов на различные мембранные рецепторы в некоторых ситуациях может вызывать синергический, но не суммарный (аддитивный) эффект. Например, холецистокинин увеличивает секрецию бикарбонатов, стимулированную секретином, но секретин не повышает секреторный ответ на действие холецистокинина.
Фазы пищеварения
Секрецию поджелудочной железы можно разделить на межпищеварительную и пищеварительную фазы. Межпищеварительная фаза заканчивается вскоре после периода интестинальной моторной активности, которую обозначают как мигрирующий миоэлектрический комплекс (ММК). ММК подразделяют на фазу I, характеризующуюся отсутствием двигательной активности, и на фазы II, III с прогрессивно усиливающейся двигательной активностью. В течение фазы I секреция ферментов и бикарбонатов поджелудочной железой, как и выделение желчи из печени и желчного пузыря, находится на самом низком уровне. В фазах II и III происходит постепенное усиление панкреатической и билиарной секреции с частичным сокращением желчного пузыря, совпадающим с повышением миоэлектрической активности. Мотилин, пищеварительный гормон, вырабатываемый в верхних отделах тонкой кишки в межпищеварительную фазу, имеет важное значение для ММК. У собак он участвует в усилении секреции поджелудочной железы в фазе III, но его роль в организме человека до конца не ясна.

Рис. 8.13. Схема стимуляции секреции белков клетками ацинуса поджелудочной железы. Сокращения; ВИП — вазоактивный интестинальный пептид; ФЛ-С — фосфолипаза С; ФИБФ — фосфатидилинозитолбифосфат; АцХ — ацетилхолин; ХЦК — холецистокинин; ИТФ — инозитолтрифосфат; gs— белок, стимулирующий присоединение гуанина; РК-А — протеинкиназа А; РК-С — протеинкиназа С; РР, РК — кальмодулинзависимый белок; ДАГ — диацилглицерол, САМ — кальмодулин.
Пищеварительная фаза секреции поджелудочной железы сложнее и разделяется на три части. Первая часть, называемая цефалической фазой (сложнорефлекторной), реализуется посредством блуждающего нерва. Эта фаза начинается с сенсорного восприятия пищи (зрительная, осязательная, обонятельная и вкусовая оценка продуктов питания). Она необходима для значительного повышения секреции ферментов и бикарбонатов. Изучение физиологии этой фазы проводилось в опытах с мнимым кормлением. В этих опытах сохранялось зрительное, обонятельное и вкусовое восприятие пищи, но пища не проглатывалась. Выявлено, что повышение панкреатической секреции при этом может быть вызвано прямым холинергетическим воздействием блуждающего нерва на ацинарные клетки, а также ацидификацией содержимого (секрета) двенадцатиперстной кишки, обусловленной повышением секреции желудочной кислоты, которое сопровождает мнимое кормление. Дуоденальная ацидификация приводит к высвобождению из слизистой оболочки двенадцатиперстной кишки секретина, который стимулирует секрецию бикарбонатов, играющих роль буфера в полости кишки. Механизм обратной связи регуляторного процесса реализуется посредством забуферивания содержимого двенадцатиперстной кишки, что тормозит выделение секретина, так как ингибируется кислотная стимуляция активности секретина. Именно таким образом угнетается секреция поджелудочной железы. В поджелудочной железе находятся пептидсодержащие (пептидергические) нейроны. Есть данные, что вагусная стимуляция также может приводить к высвобождению пептидов, подобных вазоактивному интестинальному полипептиду, гастрин-лизинг пептиду, холецистокинину и энкефалинам. Наиболее вероятно, что выделяются вазоактивный интестинальный полипептид и гастрин-рилизинг пептид. Так известно, что ВИП стимулирует и ацинарные клетки (выделение ферментов), и эпителиальные клетки протоков (выделение воды, бикарбонатов).
Вторая (желудочная) фаза начинается тогда, когда пища поступает в желудок. В течение этой фазы усиливается секреция ферментов поджелудочной железой, тогда как существенного увеличения секреции воды и бикарбонатов не происходит по сравнению с таковой в сложнорефлекторную фазу. Секреция в эту фазу стимулируется афферентными волокнами блуждающего нерва, реагирующими на растяжение желудка (фундального и антрального отделов). Содержание секретина и холецистокинина в плазме увеличивается в первые 10 мин после проглатывания пищи. Эти процессы составляют так называемый ваго-вагальный холинергический рефлекс.
Заключительная фаза пищеварения, именуемая интестинальной (тонкокишечной), завершается после поступления химуса в двенадцатиперстную кишку. Химус образуется в результате перемалывания, смешивания и сепарации проглоченной пищи. В этой фазе нейрогуморальные медиаторы способствуют более интенсивной секреции ферментов, чем во все другие фазы пищеварения. Секреция воды и бикарбонатов в этой фазе обеспечивается ацидификацией двенадцатиперстной кишки, чему также способствуют желчные и жирные кислоты. Секретин, по-видимому, является основным медиатором реакции на дуоденальную ацидификацию, но в этом процессе также имеют значение холецистокинин и холинергические влияния. Секреция ферментов в процессе интестинальной фазы стимулируется присутствием в двенадцатиперстной кишке жирных кислот, имеющих, по крайней мере, 8 атомов углерода, моноглицеридов, белков, аминокислот, кальция. Продукты переваривания углеводов играют в этом процессе небольшую роль. Помимо жирных кислот, белков, аминокислот, важное значение для полноценного, стимулированного пищей выделения ферментов имеет ваго-вагальный рефлекс. Ваготомия и введение атропина сопровождаются понижением секреции ферментов в ответ на небольшую нагрузку аминокислотами и жирными кислотами. Напротив, значительная нагрузка этими веществами потенцирует стимулы для секреции ферментов, несмотря на разрыв ваго-вагального рефлекса, и реализуется через стимуляцию выделения холецистокинина в верхних отделах тонкой кишки.
Холецистокинин-рилизинг пептид (ХРП) секретируется энтероцитами, которые неактивны в базальном, или межпищеварительном, периоде. Он необходим для стимуляции секреции холецистокинина. В межпищеварительном периоде этот пептид инактивируется под действием трипсина, содержащегося в полости кишечника. После приема пищи основное количество трипсина направлено на белки, поступающие в двенадцатиперстную кишку, поэтому ХРП в меньшей степени разрушается и в большей степени стимулирует высвобождение холецистокинина энтероцитами, а значит, и последующую стимуляцию ферментов поджелудочной железы. Таким образом, ХРП «отслеживает» готовность двенадцатиперстной кишки к перевариванию белков, способствует увеличению панкреатической секреции и улучшению управления процессом переваривания пищи. Подобный пептид имеется в соке поджелудочной железы, но в нем может также быть секретин-рилизинг пептид, высвобождаемый энтероцитами с аналогичной функцией.
Таким образом, ацидификация двенадцатиперстной кишки во всех фазах пищеварения и панкреатической секреции стимулирует выделение секретина, и этот процесс усиливается в двенадцатиперстной кишке в присутствии желчи, продуктов переваривания белков и жиров. Секретин способствует выделению бикарбонатов и воды. Холецистокинин, высвобождающийся в ответ на появление продуктов переваривания белков и жиров в двенадцатиперстной кишке, стимулирует секрецию ферментов поджелудочной железы. Выделение холецистокинина происходит, главным образом, в желудочную и интестинальную фазы пищеварительного периода. Ваго-вагальный рефлекс и пептидергические реакции имеют большое значение во всех трех фазах пищеварения (рис. 8.14).
Ингибиторы секреции поджелудочной железы
Различные вещества, ответственные за ингибирование панкреатической секреции, действуют по принципу обратной связи в процессе и после приема пищи.
Панкреатический полипептид представляет собой пептидный гормон, образующийся в островках Лангерганса и ингибирующий панкреатическую секрецию воды, бикарбонатов и ферментов. Концентрация этого пептида в плазме возрастает после мнимого кормления, либо после приема пищи, либо после экспериментальной ацидификации двенадцатиперстной кишки. Кроме того, секреция полипептида поджелудочной железой увеличивается при стимуляции блуждающего нерва, при действии холецистокинина, секретина, ВИП и, возможно, гастрина и гастрин-рилизиг пептида. Панкреатический полипептид может выступать как антагонист ацетилхолиновых рецепторов и способен ингибировать выделение ацетилхолина из постганглионарных нейронов поджелудочной железы; его конечный эффект проявляется на уровне ацинарных клеток.
Пептид YY высвобождается в дистальной части подвздошной кишки и в толстой кишке в ответ на пищу смешанного характера, но жиры, находящиеся в просвете кишки, в большей степени способны стимулировать его секрецию. Этот пептид уменьшает чувствительность поджелудочной железы к действию секретина и холецистокинина, возможно, за счет уменьшения секреции ацетилхолина и норадреналина и ингибирования выделения холецистокинина слизистой оболочкой двенадцатиперстной кишки.

Рис. 8.14. Суммарные пищеварительные эффекты панкреатического секрета. Представлены цефалическая, желудочная и кишечная фазы пищеварения. АцХ — ацетилхолин, Н+ — соляная кислота, S-RP — секретии-стимулирующий пептид, CCK-RP — стимулятор выделения холецистокинина, ВИП — вазоактивный интестинальный пептид.
Соматостатин ингибирует секрецию секретина дуоденальной слизистой оболочкой, а также чувствительность к секретину рецепторных полей. Его единственный эффект — снижение секреции ферментов и бикарбонатов поджелудочной железой. Соматостатин секретируется клетками слизистой оболочки желудка и кишечника, а также D-клетками островков Лангерганса. Однако только соматостатин, продуцируемый слизистой оболочкой тонкой кишки, оказывает ингибирующее действие на секрецию поджелудочной железы. Выделение соматостатина происходит при участии автономной нервной системы в ответ на поступление жиров и аминокислот с пищей.
Другие ингибиторы, входящие в состав гормонов эндокринных клеток островков Лангерганса, включают панкреатический глюкагон и панкреастатин, а также нейропептиды: кальцитонин-информационный пептид и энкефалины. Панкреатический глюкагон ингибирует секрецию поджелудочной железы, стимулированную холецистокинином, секретином или пищей. Частично в этом участвует холецистокинин. Глюкагон угнетает секрецию бикарбонатов, воды и ферментов. Панкреастатин ингибирует панкреатическую секрецию, тормозя высвобождение ацетилхолина эфферентными окончаниями блуждающего нерва. Кальцитонин-информационный пептид может проявлять свою активность через стимуляцию выделения соматостатина. Энкефалины и подобные им опиоиды снижают высвобождение секретина слизистой оболочкой двенадцатиперстной кишки и могут также ингибировать высвобождение ацетилхолина.