ЭВОЛЮЦИЯ ПРЕДСТАВЛЕНИЙ О СТРОЕНИИ МЕМБРАН
Биомембранология Болдырев А.А., Кяйвяряйнен Е.И., Илюха В.А.
Наличие мембран вокруг живых клеток было установлено более ста лет назад в работах К. Негели, который в 1855 году обнаружил, что неповрежденные клетки могут изменять свой объем при изменении осмотического давления окружающей среды. Эти исследования были продолжены Е. Овертоном, показавшим, что неполярные молекулы легче проходят через клеточную мембрану, чем полярные соединения. На основе этих наблюдений он впервые высказал предположение, что клеточная мембрана имеет липидную природу. Развитие идей о структуре мембран существенно продвинулось благодаря работам Е.Гортера и Ф.Грендела, проведенным в 1925 г. Эти авторы впервые выдвинули концепцию липидного бислоя. Эта идея возникла на основе простого эксперимента. Липиды эритроцитов экстрагировали ацетоном и затем получали из них тонкую пленку на поверхности воды.
С помощью поплавка сжимали слой липидных молекул на границе раздела вода/воздух до тех пор, пока этот слой не начинал оказывать сопротивление дальнейшему сжатию; это явление было объяснено образованием плотно упакованной мономолекулярной липидной пленки. Измерение площади, занимаемой липидами, и сравнение ее с площадью поверхности эритроцитов, из которых эти липиды были экстрагированы, дали соотношение 2:1. Отсюда был сделан вывод, что мембрана эритроцитов состоит из липидных молекул, расположенных в два слоя. По-видимому, этот вывод Е. Гортера и Ф. Грендела оказался правильным только благодаря взаимной компенсации ошибок (во-первых, экстракция ацетоном извлекает 6 не все липиды, во-вторых, они дали заниженную оценку площади поверхности эритроцитов, использовав для ее определения высушенные клетки). Однако в историческом плане эта работа имела большое значение, поскольку концепция липидного бислоя как структурной основы биологических мембран на самом деле оказалась верной. Мысль о том, что с мембранами связаны белки, высказана десятью годами позже Дж. Даниелли в связи с необходимостью объяснить явное расхождение между поверхностным натяжением на границах раздела масло/вода и мембрана/вода. Была высказана гипотеза, что мембрана состоит из двойного липидного слоя, и предположено, что белок располагается на ее поверхности – модель Даниели – Дэвсона, или модель «сэндвича» (рис. 1 А, Б).
Это была очень удачная модель, и в течение последующих 30 лет многочисленные экспериментальные данные, особенно полученные с помощью дифракции рентгеновских лучей и электронной микроскопии, полностью подтвердили ее адекватность. Основными компонентами биологической мембраны являются липид и белок, вопрос о взаимном расположении этих компонентов в мембране стал предметом многочисленных дискуссий, так как обнаружилось, что мембраны выполняют разнообразные функции.
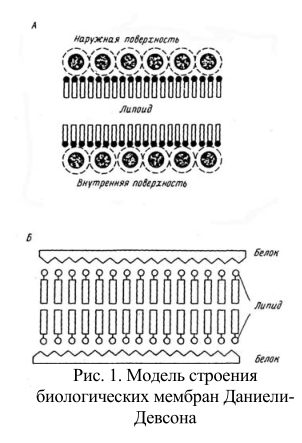
Рис. 1. Модель строения биологических мембран Даниели-Девсона
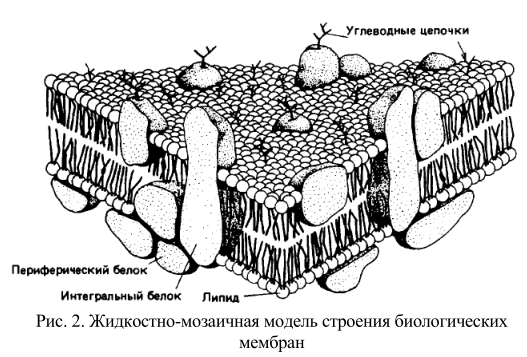
Рис. 2. Жидкостно-мозаичная модель строения биологических мембран
Быстрый прогресс в мембранологии, в результате которого сформировались современные представления, был достигнут в значительной мере благодаря успехам в изучении свойств мембранных белков. Электронно-микроскопические исследования с применением метода замораживания-скалывания показали, что в мембраны встроены глобулярные частицы. Тем временем биохимикам с помощью детергентов удалось «раздробить» мембраны до состояния функционально активных «частиц». Данные спектральных исследований указывали, что для мембранных белков характерно высокое содержание α-спиралей и что они, вероятно, образуют глобулы, а не распределены в виде монослоя на поверхности липидного бислоя. Неполярные свойства мембранных белков наводили на мысль о наличии гидрофобных контактов между белками и внутренней неполярной областью липидного бислоя. С. Сингер и Дж. Никольсон свели воедино все эти идеи, создав жидкостно-мозаичную модель. В рамках этой модели мембрана представляется как фосфолипидный бислой, в который погружены свободно диффундирующие белки (рис. 2).
Прежняя модель Даниели – Дэвсона была статичной и успешно объясняла имевшиеся в то время структурные данные, полученные с довольно низким разрешением. Начиная с 70-х гг. ХХ в. большое внимание стало уделяться изучению динамических свойств мембран и их взаимосвязи с мембранными функциями. В последние годы жидкостно-мозаичная модель (рис.2) также подверглась модификации, и этот процесс будет продолжаться в соответствии совершенствованием наших знаний. Выявляются новые функции цитоскелета. Становится ясно, что не все мембранные белки свободно диффундируют в жидком липидном бислое. Имеются данные о существовании в мембране липидных доменов. Обнаружены динамические ассоциаты липидов, обладающие более плотной упаковкой (рафты). Выявлен специфический класс амфифильных белков, которые под влиянием внеклеточных сигналов меняют свою гидрофобность и обратимо диссоциируют от мембраны. Таким образом, клеточная мембрана все более отличается по своим свойствам от «классического» липидного бислоя. Тем не менее, жидкостномозаичная модель в ее разных модификациях все еще служит в качестве концептуальной основы для объяснения многих мембранных феноменов. Сложность создания единой модели биологических мембран связана с огромным разнообразием мембранных функций.
КЛЕТОЧНЫЕ МЕМБРАННЫЕ СТРУКТУРЫ
Успехи в исследовании мембран во многом достигнуты благодаря сравнительному изучению мембран из множества разнообразных организмов. Данные, полученные при изучении клеток млекопитающих методом электронной микроскопии, свидетельствуют о наличии развитой сети внутриклеточных мембранных образований (рис. 3). Не вызывает сомнений, что основные принципы структурной организации всех мембран животной клетки по сути одинаковы. Более того, эти принципы распространяются и на мембраны растительных и бактериальных клеток.
Любая клетка имеет наружную мембрану – ее называют плазматической. Она играет роль преграды, отделяющей живое содержимое от ее неживого окружения. Но плазматическая мембрана – не просто оболочка. Она регулирует поступление молекул и ионов в клетку и выход их наружу. Кроме того, в ней находятся различные ферменты, природа которых зависит от особенностей данной клетки. Она содержит специализированные компоненты, участвующие в межклеточных контактах и взаимодействиях, в гормональном ответе и системах транспорта через мембрану как малых, так и больших молекул. Плазматическая мембрана чрезвычайно эластична, благодаря чему животные клетки могут довольно сильно изменять форму без разрыва мембран.
Большинство растительных клеток в отличие от животных не способны изменять свою форму, так как их мембраны окружены толстой, прочной и мало упругой оболочкой. Ее называют клеточной стенкой. Стенки имеются также у бактерий. Бактериальные клетки имеют довольно простую наружную оболочку, содержащую один или два слоя. Внутри клетки мембраны могут образовывать субклеточные частицы (органеллы) различного назначения. Заметим, что внешний вид органелл неодинаков в клетках разного типа.

Рис. 3. Схематическое изображение органелл эукариотических клеток животных и растений на основании данных электронной микроскопии
К ним относятся, например, митохондрии — своеобразные энергетические станции клетки, специализированные на образовании АТФ. В этих органеллах осуществляются окислительные превращения субстратов, завершающиеся образованием ATФ. В одной клетке может содержаться от нескольких десятков до нескольких тысяч митохондрий. Они сильно отличаются по размерам и очертанию. Чаще всего митохондрии имеют вид нитей или гранул (по-гречески «митос» — нить, а «хондрион» – гранула). Иногда они набухают, приобретая форму дубинки. Независимо от размера и формы каждая митохондрия содержит две мембраны – наружную и внутреннюю. Внутренняя мембрана образует складки в виде перегородок, называемых кристами, и содержит ферменты, участвующие в транспорте электронов и синтезе АТФ. Пространство, ограниченное этой мембраной, носит название матрикса, в нем протекают многие метаболические процессы. Межмембранное пространство также содержит специфические компоненты, отличающие его по составу от цитоплазмы.
Другие субклеточные органеллы – лизосомы – представляют собой окруженные мембранами органеллы, содержащие набор протеолитических и других деградационных ферментов, которые расщепляют белки, нуклеиновые кислоты, полисахариды и другие соединения. Вещества, захваченные клеткой путем эндо- или фагоцитоза, которые необходимо расщепить, доставляются в лизосомы с помощью везикул — фагосом. В лизосомах происходит также расщепление компонентов клетки, осуществляющееся в ходе клеточного цикла. Содержащиеся в лизосомах ферменты способны разрушать не только чужие вещества, но и саму клетку. Если происходит гибель клетки, мембраны лизосом разрываются и запускаются процессы автолиза. В обычных условиях лизосомальная мембрана надежно защищает клетку от воздействия собственных смертоносных ферментов.
Фагосомы – это короткоживущие внутриклеточные везикулы, образованные в результате фагоцитоза – процесса поглощения, захвата крупных частиц, комплексов, вплоть до целых клеток, например, клеток бактерий. Этот процесс характерен только для клеток некоторых типов (амебы, макрофаги). После транспорта фагосомы внутрь клетки она сливается с лизосомой, содержащей ферменты деградации.
Пероксисомы – это органеллы, которые содержат окислительные ферменты, участвующие в деградации малых молекул, таких как аминокислоты, ксантин, жирные кислоты. Их название связано с присутствием в них каталазы, которая разлагает перекиси, образующиеся в качестве продуктов окисления.
Иной вид внутриклеточных мембран образует так называемую эндоплазматическую сеть – глубокие складки, непосредственно примыкающие к плазматической мембране. Это сложная сеть цистерн и трубочек, которая занимает значительную часть внутреннего объема клетки. На мембранах шероховатого эндоплазматического ретикулума (ЭР) расположены рибосомы. Он служит местом биосинтеза белков, которые затем транспортируются к месту их функционирования. У бактерий, которые не имеют эндоплазматической сети, синтез белка, по-видимому, осуществляется на особых выступах плазматической мембраны. Области ЭР, не содержащие рибосом, называют гладким ЭР. Здесь осуществляется биосинтез стеролов, происходит десатурация (образование двойных связей) жирных кислот. Мембраны эндоплазматической сети выполняют и другую важную функцию – они обезвреживают вещества, присутствие которых нарушает нормальную работу клетки. Этот процесс называется детоксикацией. Эти процессы входят в согласованную систему транспорта электронов, осуществляющегося при участии цитохромов б5 и Р450 . Эндоплазматическая сеть клетки не однородна, а состоит из мембран, различающихся по составу и выполняемым функциям, но объединяемых в единую систему взаимодействующих друг с другом процессов.
К мембранам эндоплазматической сети примыкает так называемый аппарат Гольджи, также состоящий из ограниченных мембранами пузырьков и цистерн, собранных в стопки. Аппарат Гольджи выполняет в клетке узко специализированные, но весьма существенные задачи. Было замечено, что белки, синтезированные рибосомами, через несколько минут начинают перемещаться к аппарату Гольджи. С помощью электронного микроскопа удалось установить, что белки в аппарате Гольджи плотно упакованы в гранулы: такая упаковка белков происходит перед их секрецией. В аппарате Гольджи происходит также созревание сложных белков – например, посттрансляционная модификация гликопротеинов, синтезированных в ЭР и предназначенных для секреции, включения в плазматическую мембрану или доставки в лизосомы. Аппарат Гольджи содержит гликозидазы и гликозилтрансферазы, которые вступают в действие последовательно, по мере того как белок, подвергаемый процессингу, перемещается (вероятно, с помощью мембранных везикул) от начала аппарата Гольджи (цис-область) до его конца (транс-область). Таким образом, простые белки превращаются в сложные – гликопротеины. Прикрепление к белкам углеводных радикалов, по-видимому, облегчает их прохождение через клеточные мембраны. В некоторых клетках аппарат Гольджи формирует также лизосомы и другие субклеточные частицы. Таким образом, деятельность аппарата Гольджи связана как с построением клеточных элементов, так и с их разрушением. Соблюдение баланса между этими противоположно направленными процессами исключительно важно для жизнедеятельности клетки.
Оболочка, окружающая клеточное ядро, состоит из двух мембран, наружной и внутренней, разделенных промежутком, называемым перинуклеарным пространством. Эта мембрана происходит из эндоплазматического ретикулума и неразрывно связана с ним. Ядерная мембрана защищает святое святых клетки – хранилище ее генетической информации – от вредных внешних воздействий. Наиболее характерными морфологическим признаками ядерной мембраны являются порообразные структуры. Они имеют диаметр около 600 Å. В том месте, где расположены эти структуры, внутренняя и наружная ядерные мембраны соединяются. По краям пор наружная и внутренняя мембрана ядра сливаются в одну общую мембрану. Полагают, что поры позволяют комплексам мРНК-белок переходить из ядра в цитоплазму, а регуляторным белкам перемещаться в обратном направлении, из цитоплазмы в ядро. Таким образом, ядерная мембрана контролирует перенос информации между ядром и остальной частью клетки. Есть также сведения, что мембрана обеспечивает энергией процессы, протекающие внутри ядра.
Хлоропласты – это органеллы, содержащие фотосинтетический аппарат. Они характерны для растений и некоторых микроорганизмов. Наружная оболочка хлоропластов образуется двумя мембранами, а внутренняя область составляет строму. В строме находятся тилакоидные мембраны, где локализованы компоненты системы фотосинтеза. На отдельных участках тилакоидные мембраны плотно упакованы в стопки, а на других – обращены непосредственно к строме. Состав плотно упакованных и обращенных в строму доменов тилакоидной мембраны различен, что указывает на различие их функций.
Все клеточные мембраны представляют удивительные биологические конструкции. Они отличаются исключительной тонкостью, при этом обладают высокой прочностью на разрыв, устойчивостью и гибкостью, а по электроизоляционным свойствам превосходят многие изоляционные материалы, применяемые в технике. Общая площадь мембран в органах и тканях организма достигает огромных размеров. Печень крысы весит всего 6 г, суммарная же площадь ее клеточных мембран составляет несколько сотен квадратных метров. В эндоплазматической сети печени на каждый миллиграмм белка приходится 0,5 м 2 мембран.
БИОЛОГИЧЕСКИЕ ФУНКЦИИ МЕМБРАН
Несмотря на многообразие различных типов клеток, их мембраны выполняют общие биологические функции. Прежде всего, они отграничивают живое от неживого. Более того, они организуют внутри клетки компартменты с различными свойствами. С их помощью происходит отделение содержимого компартментов от окружающей их среды. В каждом компартменте мембраны обеспечивают сохранение специфических физико-химических условий. Поэтому по обе стороны мембраны такие условия среды как кислотность, концентрация растворенных веществ, электрический потенциал, как правило, не одинаковы.
Однако мембраны не только разделяют клетку на отдельные компартменты, но и участвуют в регуляции метаболических сигналов, которые передаются между наружной и внутренней сторонами этих компартментов. Это может проявляться в виде физического переноса ионов или молекул через мембрану или при помощи конформационных изменений, индуцируемых в мембранных компонентах. Таким образом, мембраны контролируют проникновение в клетку и выход из нее метаболитов. С помощью мембранных рецепторов они реагируют на внешние сигналы и трансформируют их, то есть способны классифицировать и избирательно модулировать их (усиливать важные и снижать до уровня шумов второстепенные), передавая внутрь клетки существенную информацию. Мембраны способны обеспечивать образование и поддержание разности потенциалов, а также транспортировать мембранный потенциал вдоль по мембранным индукторам, позволяя использовать этот специфический вид энергии в разных частях клетки.
Кроме того, с мембранами связано функционирование многих клеточных ферментов. Мембраны оказывают большое влияние на процессы, протекающие внутри клетки, изменяя их активность. Некоторые ферменты активны только тогда, когда они прикреплены к мембране; другие, наоборот, в этом состоянии не проявляют активности и начинают действовать лишь после отщепления их и выхода в цитоплазму. Поэтому важным свойством мембран является способность создавать специальную среду для защиты гидрофобных белков от водной атаки и обеспечения их функций, то есть в мембране создаются специальные условия для протекания реакций, осуществляемых гидрофобными белками. Одновременно мембранные липиды осуществляют контроль за взаимодействием между отдельными белками, погруженными в мембранную толщу. Некоторые ферменты образуют своеобразные мембранные ансамбли, которые осуществляют цепь последовательных превращений именно благодаря тому, что их компоненты объединены общностью локализации, организованы мембраной. Благодаря этому обстоятельству повышается эффективность суммарного процесса. Имеются ферменты, которые, действуя на мембраносвязанные субстраты, участвуют тем самым в биосинтезе мембран.
С участием мембран в той или иной степени осуществляется большинство жизненно важных функций, например, протекают такие разные процессы, как репликация прокариотической ДНК, биосинтез белков и их секреция, биоэнергетические превращения, а также функционирование систем гормонального ответа. Важная сторона ферментативной деятельности мембран связана с координацией множества химических реакций, протекающих в клетке. Для этого мембраны объединяют различные ферменты в единый конвейер, в котором каждый фермент действует в строгом соответствии с остальными.
Мембраны участвуют во взаимодействии клеток со средой. Это свойство лежит в основе обеспечения специфики межклеточных контактов и иммунологических ответов. Клетки узнают себе подобных, вступают с ними в контакт, передают разнообразную информацию. Если измельчить эмбрионы амфибии до состояния свободных клеток и перемешать их, можно наблюдать, что через некоторое время клетки самопроизвольно начинают «сортироваться»: родственные клетки объединяются в пласты, дающие начало тканям, и, в конце концов, вновь образуются структуры, напоминающие эмбрион. Эта поразительная способность клеток, несомненно, зависит от свойств наружных клеточных мембран, их способности узнавать себе подобных. Благодаря этому свойству клетки способны создавать ткани, органы и сами организмы.
Большинство мембран, кроме этих общих функций, выполняют и специальные функции. Например, мембраны митохондрий и хлоропластов зеленых растений осуществляют трансформацию энергии. Мембраны, расположенные в стенках кишечника, выполняют функции, связанные с процессами пристеночного пищеварения. Мембраны нервных клеток генерируют электрические импульсы. Некоторые клетки, например, палочки сетчатки глаза, имеют высокоспециализированные мембраны, позволяющие выполнять уникальные функции. Мембраны мышечных клеток участвуют в инициации и регуляции сокращения. Клетки органов чувств содержат специализированные мембраны, преобразующие энергию света и звука в электрические импульсы и передающие центральной нервной системе информацию о запахах, изменениях температуры и давления.